
Parasitic Diseases Journal of The Official Organ of the Indian Society for Parasitology
ISSN: 0971-7196 Journal of Volume 30 Number 1 June 2006 (Released August 2006) Parasitic Diseases C SO IET Y FOR PA R THE IND LOGY ITO H kkj Published by AS I AN The Official Organ of the Indian Society for Parasitology r h; 1973 a?k i j t ho h f o K k u l The Indian Society for Parasitology Electronic version available on ISP Website www.parasitologyindia.org JOURNAL OF PARASITIC DISEASES (ISSN: 0971-7196) Editor-in-Chief Professor Prati Pal Singh National Institute of Pharmaceutical Education and Research Sector-67, Phase-X S. A. S. Nagar-160 062, India Managing Editor Assistant Managing Editor Dr. Varsha Gupta Deptt. of Microbiology Govt. Medical College & Hospital Chandigarh-160 032 Dr. Savita Singh National Institute of Pharmaceutical Education and Research S. A. S. Nagar-160 062 Advisory Board Dr. A. B. Chaudhary, Kolkata Prof. R. C. Mahajan, Chandigarh Dr. G. P. Dutta, Lucknow Dr. V. P. Sharma, New Delhi Prof. N. K. Ganguly, New Delhi Editorial Board Prof. M. C. Agrawal, Jabalpur Prof. Irfan Ahmed, Aligarh Dr. H. K. Bajaj, Hissar Prof. H. S. Banyal, Shimla Prof. Neelima Gupta, Bareilley Prof. B. C. Harinath, Sevagram Dr. D. C. Kaushal, Lucknow Dr. S. L. Hoti, Pondicherry Prof. R. Madhubala, New Delhi Prof. Sandeep Malhotra, Allahabad Prof. Nancy Malla, Chandigarh Dr. J. R. Rao, Izatnagar Prof. M. L. Sood, Ludhiana Dr. B. K. Tyagi, Mdurai Journal of Parasitic Diseases is published biannually by the Indian Society for Parasitology in June and December in each calender year. The subscription price for libraries and other multi-reader organizations for each number is Rs. 400 in India and US $ 100 elsewhere. Subscription by Demand Draft in favour of 'The Indian Society for Parasitology' should be sent to Dr. J. K. Saxena, Secretary, The Indian Society for Parasitology, Division of Biochemistry, Central Drug Research Institute, Chattar Manzil, Lucknow - 226 001, India. JOURNAL OF PARASITIC DISEASES Volume 30, Number 1, June 2006 The official organ of THE INDIAN SOCIETY FOR PARASITOLOGY Central Drug Research Institute, Lucknow-226 001, India Editorial Office: National Institute of Pharmaceutical Education and Research Sector-67, Phase-X, S. A. S. Nagar-160 062, India Phone: 0172-2214682-87; Fax: 0172-2214692; E-mail: drppsinghjpd@yahoo.co.in Journal of Parasitic Diseases Copyright © 2006 The Indian Society for Parasitology All Rights Reserved No part of this publication may be reproduced or utilized in any form or by any means, electronic or mechanical including photocopying, recording or any other information storage and retrieval system without the permission in writing from copyright owner. JOURNAL OF PARASITIC DISEASES Volume 30 Number 1 June 2006 CONTENTS A Note from the New Editor-in-Chief 1-3 Reviews Histochemical, biochemical and immunological studies in Haemonchus contortus (Nematoda: Trichostrongyloidea) - an Indian perspective. M. L. Sood Immunological perspectives and malaria vaccine. H. S. Banyal and N. Elangbam 4-15 16-29 Original papers Isolation and characterization of the paraflagellar rod proteins of Leishmania donovani. A. Lahiri and A. Bhattacharya 30-36 Onchocercosis in Benue state, Nigeria: comparative epidemiological amongst the Etulo and Idoma ethnic groups. E. A. Omudu and B. O. Atu studies 37-40 Distribution of iron in plasma, erythrocytes and tissues of calves with the progression of Theileria annulata infection. N. Sangwan and A. K. Sangwan 41-44 Mosquito breeding in riceland agro-ecosystem near Chennai, Tamil Nadu, India. J. Ravindran and J. Williams 45-52 Random amplified polymorphic DNA of Trichomonas vaginalis isolates from Tarbiz, Iran. R. Jamali, B. Zareikar, A. Kazemi, M. Asgharzadeh, S. Yousefee, R. Estakhri, S. Montazer and A. Ghazanchaei 53-57 Two new species of Trypanosoma from freshwater fish (Heteropneustes fossilis and Channa punctatus) from Bareilley, India. D. K. Gupta, N. Gupta and R. Gangwar 58-63 Impact of anthelmintic therapy on live weight gain in gastrointestinal nematodeinfected goats. A. K. Jayraw and Y. V. Raote 64-67 Ultrastructure, differential density and distribution pattern of polymorphic microtriches in tegument of Stilesia globipunctata infecting Ovis aries (sheep). C. Venkatesh, K. Ramalingam and V. Vijayalakshmi 68-75 The protozoan fauna living in the digestive system of Periplaneta americana in Kolkata, West Bengal, India. J. Ghosh and A. Gayen 76-80 Chelatrema neilgherriensis n. sp. (Trematoda: Gorgoderidae) infecting the freshwater fishes from Noolpuzha river in Wynad district, Kerala, India. K. T. Manjula and K. P. Janardanan 81-84 Short communications Haemato-biochemical studies on fowl coccidiosis in layer birds. N. D. Hirani, J. J. Hasnani, R. S. Joshi and K. S. Prajapati 85-88 Re-redescription of Dissurus farrukhabadi Verma, 1936 (Digenea Echinostomatidae) with a discussion of the genus Dissurus Verma, 1936. P. C. Gupta and R. B. Singh 89-91 A case of vaginal bleeding due to leech bite. R. P. Ganguly, M. S. Mukhopadhyay and K. K. Patra 92-93 Field evaluation of a rapid immunochromatographic test kit for the diagnosis of Plasmodium falciparum and non-falciparum malaria parasites from Sonpur district, Assam. C. Rajendran and S. N. Dube 94-97 Journal of Parasitic Diseases: Vol. 30, No. 1, June 2006, 1-3 J PD A Note from the New Editor-in-Chief “What the Author Expects from the Editors”. This has been expressed by Dr. Earl H. Wood of Mayo Clinic as “I expect the editor to accept all my papers, accept them as they are submitted, and publish them promptly. I also expect him or her to scrutinize all other papers with utmost care, especially those of my competitors”. My association with Journal of Parasitic Diseases (started as Indian Journal of Parasitology) is both personal and somewhat historical, and goes nearly 30 years back when in 1976 the first issue of the journal was released in the main conference hall of the Central Drug Research Institute, Lucknow. Certainly, it must have been born out of the hard work and uncompromised dedication of several Members of the Society, and the vision of its Founder Editor-in-Chief, late Dr. B. N. Singh. And with this issue, in a new style and format, my tenure as Editor-in-Chief has just begun. The Editor-in-Chief of a journal is ultimately responsible for its quality standards and acceptability by the audience it addresses. His or her singular goal should be to deliver the readers a package of good science in a good and simple language. Therefore, the final decision to accept or not to accept a manuscript for publication or to send it back for modification must be the responsibility of the Editor-in-Chief only. For arriving at such a decision, he or she should carefully go through the manuscript and take into consideration the recommendations, and the reports and comments of the Editorial Board Members and Reviewers/ Referees/Consultants, respectively. However, for the resolution of complicated and controversial matters, the Editor-in-Chief should also be willing to take the help of Advisory Board Members, should such a situation arise. The Members of Editorial Board, who must be experts in their respective fields, in turn, have a very important responsibility of sending the manuscripts to suitable Reviewers/Referees/Consultants, determine the quality of these manuscripts and then send their recommendations to the Editor-in-Chief. Their recommendations regarding the suitability of the manuscripts for publication, revision or rejection are crucial for the Editor-in-Chief in making a final decision, and thus, in maintaining the quality standards of the journal. Therefore, the Editor-inChief must decide upon an appropriate size of the Editorial Board taking into consideration the expertise of its Members which should be in consonance with both the dimensions of the scope of the journal and the number of manuscripts to be handled. The Reviewers/Referees/Consultants read, evaluate and return the manuscripts along with their comments prepared in accordance with the instructions provided. These comments must be specific and not abstract, point errors related both to fact and interpretations, indicate inaccuracies and ambiguities, and also must clearly suggest as to how to condense, enlarge and improve upon the style of writing. In biological sciences, hardly 5% manuscripts are accepted without any revision. “The Guidelines for Reviewers, American Society for Microbiology”, formulated based on the policies recommended by a committee of the Council of Biology Editors is usually very helpful for the Reviewers/Referees/Consultants. The help provided by them is generally acknowledged in the form of listing their names in the last issue of the year. Because a manuscript is the intellectual property of its authors, it must be treated as a privileged confidential communication throughout, till published. The Managing Editor of a journal is usually not 2 involved in decisions related to the acceptance or nonacceptance of a manuscript. Rather, he or she is supposed to provide the Editor-in-Chief various support services during the review process. However, the main function of a Managing Editor starts only after a manuscript has been accepted for publication, and he or she takes the responsibility to convert it into a printed product i. e. a published paper. The Copy Editors and Production Editors, usually associated with big journals are responsible for the final product as related to grammar, spellings, syntax, style polishing etc., and the quality of page layout and image resolution, respectively. Therefore, in my opinion, the entire Editorial Board must meet at least once during the year, and our annual meetings can be the most appropriate occasions for such meetings. It is aptly said that the reputation of a journal primarily depends upon the quality of research papers it publishes. I shall, therefore, with the help of Advisors, Members of the Editorial Board and Reviewers/ Referees/Consultants, strive hard to take the standards of our journal to new heights. Towards this end, I shall expand the current Editorial Board, both in terms of the number of its Members and the diversity of their fields of specialization to encompass various aspects of parasitology. The speed of the editorial processing of the manuscripts submitted for publication leading to its final acceptance or non-acceptance is also important. I shall try to strictly follow the 15-day duration for the Reviewers/ Referees/Consultants to submit their reports. To the extent possible, I shall encourage the use of electronic mail for sending the manuscripts for review and for receiving the review reports. This can be expected to reduce both time and cost. The job of an Editor-in-Chief is a very responsible one. I would, therefore, be a remiss if I did not acknowledge the attendant challenges that I expect to face. First, I believe, is to implement again the very vision with which the journal was conceived and started. There is no doubt that the primary constituency of Journal of Parasitic Diseases is basic parasitology. During my informal discussions with various members of the Society, I gathered the impression that a large number of them feel that the journal has seriously departed from its primary constituency. Certainly, such wheels must have turned slowly over a period of time. I would, therefore, with the help of our authors/contributors and Editorial Board Members, like to improve upon this state and try to move the journal back to basic Prati Pal Singh parasitology. This certainly does not mean that the newer cellular and molecular, and other important and emerging dimensions of parasitology shall be neglected. Second, the journal should orient itself in such a way, that it must make an impact on the conduct of the theory and practice (including clinical) of parasiotology in our country. This may not turn out to be an easy task. The third, I feel is that there are several areas in parasitology which continue to remain controversial, conflicting, confusing and conjecturing. Therefore, Journal of Parasitic Diseases must specifically publish comprehensive and accessible reviews which should try to focus on these areas of parasitology. Towards this end, I intend to invite manuscripts from experts who would be willing to specifically contribute such reviews. However, this notwithstanding, the journal intends to publish at least two invited reviews on various important aspects of parasitology in its each issue. Parasitology research is a very big field spanning mainly into areas like agricultural, veterinary, medical and general parasitology. These areas, in turn, have their own sub-areas and allied areas. Put together, they all constitute a big world of parasitology. In our country Journal of Parasitic Diseases is probably the only journal that is specifically devoted to parasites and the diseases caused by them. Therefore, it should be the aim of the journal to adequately address all such areas and sub-areas effectively. Additionally, Journal of Parasitic Diseases should also try to address to areas related to parasitology and community/society, parasitology and economics, and parasitology and nation development. One of the most important policies that I have already introduced is the “Tutorial Editorial Policy”. This policy aims towards working of the Editorial Board with the authors, to improve the quality of their submitted manuscripts so as to make them publishable. Experience has shown that a greater and intense interaction between the authors and Editorial Board has always been beneficial in improving both the quality of science and of writing. Scientific writings are different from any other type of writings and at times not easy, and thus require considerable practice. Unfortunately, in our country, unlike in several other countries, most of the universities do not have a formal course related specifically to scientific writings. It is now well known that in many reputed journals, mostly the manuscripts are not turned down because of bad science but bad writing. The effects of Editor's Note this policy may not become visible very soon, but certainly in the long run it is going to be beneficial, both to the authors and the journal. I would like to take this opportunity to assure our authors that the entire Editorial Board is on their side, to help them in publishing good science. Here, I must bring to your kind attention that our journal has a big problem of not having a suitable number of quality manuscripts available for publication. And, this often results in the delayed release of a particular issue of the journal. In an editorial titled “On the Future of Scholarly Journals”, published in Science (17 April 1998, 279, 359), the author Alen M. Edelson has very appropriately emphasized the potential impact of digital/electronic publishing on print publishing. Presently, most of our print journals are suffering from financial crunch because of declining library budgets, escalating costs, reduction/cancellation in subscriptions and, of course, increasing manuscript pressure. Digital/electronic publishing, on the other hand, can be expected to be cost-effective and would allow greater and relatively more convenient accessibility to scientific information. Therefore, I believe in the need that Journal of Parasitic Diseases also must run with the times, and should evolve strategies to digitalize it in a bigger way, including online submission and processing of manuscripts. Nevertheless, it is becoming increasingly clear that electronic journals will also be fraught with their own 3 sets of problems, and may not be able to make print journals obsolete so soon. In closing, I would like to sincerely thank Dr. V. P. Sharma (Chairman) and Members of the committee to constitute new Editorial Board Prof. M. S. Jairajpuri (former President of the Society), Prof. Veena Tandon (President of the Society), Prof. Nancy Malla (former Editor-in-Chief), Dr. J. K. Saxena (Secretary of the Society), Dr. L. M. Tripathi (Treasurer of the Society), Dr. S. Dutta and Dr. S. L. Hoti, and to the Society at large, for the honour of offering me this important responsibility and for reposing confidence in me. I should like to take this opportunity to especially thank Prof. R. C. Mahajan and Dr. G. P. Dutta for their valuable suggestions. I should also like to thank the authors of this issue and to the future authors. I deem it my greatest pleasure to thank Prof. P. Ramarao, Director, National Institute of Pharmaceutical Education and Research, for his help and encouragement without which it would not have been possible for me to undertake this responsibility. Finally, I thank our printer Mr. Lalit Azad, for his keen interest and excellent work in a timely manner. I am confident that with our joint efforts, Journal of Parasitic Diseases will continue to maintain its high standards and surge to new heights. I shall continue to interact with you all from time to time. Prati Pal Singh, Ph. D., F. N. A. Sc. Editor-in-Chief Review Journal of Parasitic Diseases: June 2006, Vol. 30, No. 1, 4–15 J PD Histochemical, biochemical and immunological studies in Haemonchus contortus (Nematoda: Trichostrongyloidea) - an Indian perspective M. L. Sood Department of Zoology, Punjab Agricultural University, Ludhiana. ABSTRACT. Haemonchus contortus is the most pathogenic nematode parasite of sheep/goats in tropics/subtropics. Presently, the different aspects of H. contortus research in histochemistry [absorptive surfaces-structure and composition (the body wall, the gut), anthelmintic effects; spicules and gubernaculum], biochemistry [inorganic elements; carbohydrates (glucose and glycogen utilization, glycolysis, TCA cycle, pentose phosphate pathway]; amino acids (composition, metabolism, anthelmintic effects); lipids (composition, metabolism, enzymes, anthelmintic effects), proteins; nucleic acids; nutrition (digestive enzymes), biochemistry in taxonomy; miscellaneous] and immunology have been reviewed in Indian context. Key words: biochemistry, Haemonchus contortus, histochemistry, immunology, India INTRODUCTION Heamonchus contortus (Rud.,1803) commonly known as the twisted stomach worm, is a bloodsucking nematode parasite, primarily occurring in the abomasum (Fig.1) of small ruminants, notably sheep and goats. It has been ranked as the most important parasite of small ruminants in all regions across the tropics/subtropics (Anonymous, 1992). Haemonchosis, the disease caused by this nematode is responsible for considerable economic losses worldwide. In hyperacute disease, death of the host may occur within one week without significant signs (Fraser, 1991). Over the years, attempts have been made to study various aspects of Haemonchus spp. from different parts of the world (see Sood and Kapur, 1982a). Earlier, Haemonchus research in India was reviewed Fig. 1. A part of goat abomasum cut open to show Haemonchus contortus, in situ. Inset, adult female and male (From Sood and Kapur, 1982; with permission from Shidha Publications, Ludhiana, India). Corresponding author: Prof. M. L. Sood, 500/4, Model Town (Club lane), Ludhiana - 141 002, Punjab, India. E-mail : mlsood@rediffmail.com (Sood, 1981) under the titles : morphology and d i s t r i b u t i o n , b i o c h e m i s t r y, i m m u n o l o g y, development and cytology, life-history, pathogenesis, Haemonchus histochemistry, biochemistry and immunology 5 clinical symptoms, diagnosis, epidemiology, treatment and prophylaxis. Subsequently, in a review on haemonchosis in India, Sood (2003) covered epidemiology, pathology, diagnosis and control, including resistant strains. Presently, the upcoming areas of Haemonchus research, and the ones not fully covered in the above reviews have been taken up. Hopefully, these three reviews, covering separately the various aspects, would provide an up-dated consolidated account of 'Haemonchus to haemonchosis' in India. the cuticle (Sood and Kaur, 1977), has been well documented (Sood and Kaur, 1976). HISTOCHEMISTRY Histochemical studies have been made on the absorptive surfaces (including in vitro anthelmintic effects) and of the spicules and gubernaculum. Absorptive surfaces Parasitic nematodes have two absorptive surfaces- the external surface or the body wall and the gut, though latter is the main functional unit from nutrition point of view. The body wall in nematodes is composed of the cuticle, an underlying cell layer (hypodermis or epidermis), and the longitudinally orientated somatic musculature (Wright, 1987). Nematode cuticle is one of the most complex acellular structures synthesized by a living organism. In order to understand the transport of solutes in the body wall, experimental determination of diffusion coefficients of NaCl and KCl in adult H. contortus at 298. 16 k has been made (Sood et al., 1999). The nematode gut consists of three parts, a muscular, cuticular lined pharynx (oesophagus), a relatively straight intestine, whose wall is one cell thick and a rectum or cloaca, which like pharynx is lined by cuticle (Bird, 1971; Chitwood and Chitwood, 1974). Nematodes do not have a continuous muscle sheet around the intestine and the pseudocoelomic fluid is in direct contact with the basement membrane. The nematode intestine is, therefore, an ideal cell monolayer to study the transport mechanisms, the only diffusion barrier being the basement membrane (Barrett, 1981). Absorptive surfaces in H. contortus have been reviewed (Sood, 1999), under the titles: structure and composition, immunology and anthelmintic effects. Structure and composition The body wall: The morphology of the vulvar configurations in the female H. contortus, formed by The histochemical studies on the body wall of H. contortus (Sood and Kalra, 1977) have revealed that the cuticle is mainly proteinaceous in nature. The lipids and PAS-positive materials are only present in cortical layers. In addition, haemoglobin and acid phosphatase (AcPase) are also present. Glycogen, lipids, RNA, AcPase and alkaline phosphatase (AkPase) have been reported in the hypodermis. The oval dense body is composed of keratinous and collagenous proteins associated with acid mucopolysaccharides. Muscles carry a greater concentration of glycogen granules and phospholipids. The functional significance of these components has been fully discussed. The gut: Singh and Johl (2001) studied the structure of the fore-gut (stomodaeum). Histomorphology of the intestine has also been studied (Johl, 2003). Sood and Sehajpal (1978) made morphological, histochemical and biochemical studies. The intestinal epithelium (IE) is provided with a well-developed brush border (BB) which contains periodic acidSchiff- positive mucoproteins. The IE stores glycogen and lipids. It stains diffusely for phospholipids and general proteins and also for terminal-NH2 group. The presence of Fe2+ and Fe3+ containing pigments and activities of AcPase and AkPase, glucose-6phosphatase (G-6-Pase) and 5'-nucleotidase have been abserved in IE. Biochemically, pH optimum for intestinal AcPase has been found to be 4.8. BB shows positive reactions for Acpase and G-6-Pase, and negative for AkPase and 5'-nucleotidase. Presence of enzymes in the BB is related to extracellular digestion and absorption of nutrients. Anthelmintic effects Study of alterations induced in the absorptive surfaces (and neuromuscular system) of the parasites by the in vitro incubations with the anthelmintic drugs is of prime importance, particularly in view of the repeatedly reported resistant strains of H. contortus. The mechanisms by which the drugs act on these absorptive surfaces are quite obscure. During their absorption, anthelmintics are expected to induce some cellular and chemical changes in the absorptive surfaces. Anthelmintics during their absorption affect/and/or modify the enzyme activity and may also alter the normal metabolism of the absorptive surfaces. 6 Sood Anthelmintic effects (and other aspects) on the absorptive surfaces have been reviewed (Kaur and Sood, 1986). well as neuromuscular system) and hypodermis emphasize that both absorption and motility of the parasite are effected. The morphological and histochemical effects of in vitro incubations with thiabendazole, morantel tartrate, tetramisole hydrochloride and piperazine hexahydrate on the body wall and intestine have been investigated (Sood and Kaur, 1982). Similar effects of dl-tetramisole (TMS) and rafoxanide (RFX) have been observed for AkPase, AcPase adenosine triphosphatase (ATPase) and G-6-Pase (Kaur and Sood, 1982 d). Also, histological study on the effects of these two anthelmintics has been made (Kaur and Sood, 1983a). In vitro alterations induced by Nilzan (NLZ) and albendazole (ABZ) have also been studied in AcPase, AkPase, ATPase, G-6-Pase, cytochrome oxidase, monoamine oxidase, non-specific esterases, acetylcholine esterase (AChE), and in succinic, lactic, glutamate and glucose-6-phosphate dehydrogenases, reduced nicotinamide, adenine dinucleotide diaphorase and reduced nicotinamide adenine dinucleotide diaphorase (NADH-D) and reduced nicotinamide adenine dinucleotide phosphate diaphorase (Kaur and Sood, 1990). Also, the effects of Nilverm (NLV) and Nilverm forte (NLF) (Kaur and Sood, 1992a) and thiophenate (TP) and fenbendazole (FBZ) (Kaur and Sood, 1992b) have been abserved for these enzymes except NADH-D. Also, the morphological and histochemical effects of ALB, FBZ, TP, TMS, dl-tetramisole and oxyclozanide (TO), and levamisole HCl (LMS) have been studied on the absorptive surfaces (and neuromuscular system) of adult H. contortus. ABZ, FBZ, TP, TO, HCl and LMS reduced the quantity of neutral mucopolysaccharides in the intestine. TO and ABZ caused the loss of acid mucopolysaccharides in the microvilli of intestine. Loss of lipids from the intestine was evident after TO, ABZ, FBZ and TP treatments. LMS caused accumulation of very large lipid droplets in the intestine (Kaur and Sood, 1996). Spicules and gubernaculum In general, morphological changes are most evident in the intestine and muscles. Enzymatic intensities and alterations induced by the in vitro incubations of the drugs in H. contortus are stronger in the intestine than in any structure, indicating it to be the main structure involved in the absorption and also in the action of various anthelmintics. Different structures of the parasite respond differently to a particular drug, indicating the response and function correlation. The alterations induced by the drugs in the intestine (as Morphological and histochemical studies on the spicules and gubernaculum (Sood and Kaur, 1983), have revealed that these are mainly proteinaceous. The sclerotized part consists mainly of keratin and collagen. The presence of traces of AcPase in the spicules may represent some metabolic activity. The protoplasmic part is composed of proteins, carbohydrates and lipids. BIOCHEMISTRY Much of the interest in parasite biochemistry comes from the ways in which the metabolic pathways have been modified to suit the highly specialized parasitic mode of life. In addition to intrinsic interest, parasite biochemistry has great practical importance through chemotherapy and vaccine production, and in understanding of the complex association involved in the host-parasite relationship. However, information in parasite biochemistry is patchy, and mainly the large-sized worms have been exploited (Barrett, 1981). Of the various biochemical parameters, in H. contortus, carbohydrates and lipids have been studied more extensively compared to others. Further, most of these studies are restricted to adult form, though in L3 carbohydrate catabolism has been studied in detail. Moreover, variation in metabolic pathways due to strains poses a great problem. Biochemistry of Haemonchus has been reviewed (Kapur and Sood, 1987a) under the titles: chemical composition (carbohydrates, lipids, proteins, amino acids, nucleic acids, hormones, inorganic elements, pigments), absorption/transport, biosynthesis (carbohydrates, lipids, proteins, amino acids, nucleic acids), catabolism/utilization (carbohydrates-glycolysis, TCA cycle), carbon dioxide fixation, electron transport system, pentose phosphate pathway, glyoxylate cycle, strain variations in energy metabolism; lipids; proteins; biochemistry in taxonomy and miscellaneous. Inorganic elements These play a significant role in the physiology of parasites. Their impotance is also demonstrated by the influence of mineral deficiencies in host's diet. Haemonchus histochemistry, biochemistry and immunology 7 Various elements detected in H. contortus (Sood and Kapur, 1980) include phosphorous (P), zinc (Zn), calcium (Ca), iron (Fe), magnesium (Mg), copper (Cu), manganese (Mn), boron (B) and potassium (K), in decreasing concentrations in female. In the male also, same elements are present, with the exception of B. The relative order of occurrence in the male is P>Ca>Zn>Fe>Mn>Mg>Cu>K. Presence of more Ca, P and Fe in male has been discussed. of glycogen utilization is similar both in male and female (Premvati and Chopra, 1979). Under in vitro conditions, the worms readily utilize glucose from the medium (Chopra and Premvati, 1977; Kaur and Sood, 1982a). At the same time, glucose is excreted and this excretion is possibly at the expense of endogenous glycogen reserves, which falls rapidly with time. An outstanding feature of carbohydrate catabolism in nematodes is the production of reduced organic end products, even under aerobic conditions. Carbohydrates Carbohydrates form the chief energy source in parasitic nematodes. In view of the importance of carbohydrates in helminths, any difference in their carbohydrate metabolism and that of their hosts might be usefully exploited in helminth control. The outstanding feature of carbohydrate breakdown in nematodes is the production of reduced organic end products and this occurs even under aerobic conditions. Carbohydrate metabolism in H. contortus has been reviewed (Kaur and Sood, 1983b) under the titles : glucose and glycogen utilization, carbohydrate metabolism, including the glycolytic pathway, the pathway of CO2 fixation, the TCA cycle and the pentose phosphate pathway. Glucose and glycogen utilization: Glucose in very important energy source for many helminths inhabiting the gut of vertebrates. It is generally believed that helminths absorb glucose against a concentration gradient and use their endogenous carbohydrates as an energy source only when it is unobtainable from outside. Similarly, glycogen in most of the nematodes provides a significant reserve store of energy, particularly in forms which are parasitic in animals and which exist in environments of low oxygen tension. Amount of glucose and glycogen contents in H. contortus has been determined. It contains considerable amount(8-12% fresh tissue of glycogen; Chopra and Premvati, 1977; Premvati and Chopra, 1979; Kaur and Sood, 1982a). Famale has more glycogen than male (Premvati and Chopra, 1979). This is possibly related to the reproductive role of female rather than with general metabolism. Adult worm utilizes glycogen reserves rapidly. In male, it decreases less rapidly with time, while lin female, it decreases exponentially with time. Thus, in female, rate of glycogen utilization at any time is directly proportional to glycogen content. However, initial rate The in vitro effects of TMS and RFX on these constituents have also been studied (Kaur and Sood, 1982a). The pathway of carbohydrate breakdown in H. contortus has been worked out as follows: Glycolysis: Adult worm utilizes glucose both aerobically and anaerobically. Under both aerobic and anaerobic conditions, end products of glucose metabolism include CO2, propanol, acetate, and npropionate as the major and ethonol, lactate and succinate as the minor components. It is unique among nematodes that propanol is a major excretory product. This is possibly produced by reduction of propionate and hence glucose catabolism in H. contortus does not indicate a major departure from the known pathways of anaerobic glucose utilization. Production of lactate from glucose in H. contortus has been demonstrated by Chopra and Premvati (1977). In female 90-100% of glucose and in male 80-90% is catabolized to lactate. Both glucose consumption and lactate production decrease with the progression of incubation. Enzymes of glycolysis have been detected in adult H. contortus (Kaur and Sood, 1982b). These include hexokinase, phosphoglucomutase, phosphoglucoisomerase, aldolase, glycerladehyde-3phosphate dehydrogenase, phosphoglycerate kinase, phosphoglyceromutase-enolase-pyruvate kinase, pyruvate kinase (PK), and lactate dehydrogenase (LDH). Low PK and LDH activities suggested an alternate pathway from phosphoenolpyruvate. LDH exhibits optimum activity (about 180 nmoles/min/mg protein at a pH of 6.6 and a temperature of 37° C (Kapur et al., 1985). Thermostability of LDH has also been studied (Harpreet et al., 1991). Varying degrees of inhibition of glycolytic enzymes have been observed with 50 µg/ml of TMS and RFX treatments, former being more effective. These effects may block the glycolytic pathway and deprive the 8 parasite of its ATP production (Kaur and Sood, 1982b). TCA cycle: Studies on aerobic catabolism of carbohydrates in helminthology is an important area of research. In small nematodes, the TCA cycle plays an important but not exclusive part in their energy economy (Ward, 1982). The occurrence of TCA cycle in parasites has not been demonstrated with the same precision as in vertebrate tissue or bacteria. A functional TCA cycle may exist in the developing eggs and larvae of H. contortus. The presence of aerobic enzyme systems in the larvae may be a preparation for the next stage of development, since development of larvae to adults requires oxygen. Various enzymes of the TCA cycle viz. aconitase (ACO), isocitrate dehydrogenase (ICDH), succinate dehydrogenase (SDH), fumerate reductase (FR), fumarase and malate dehydrogenase (MDH) have been detected in adult H. contortus (Kaur and Sood, 1983c). Low activities of ACO and ICDH suggest that TCA cycle has a minor function and the pathway of CO2 fixation is the major pathway in the energy metabolism of the parasite. Incorporation of carbon into proteins and carbohydrates (Kapur and Sood, 1986c). and of C14 into various amino acids (Kapur and Sood, 1984b), also give evidence for the TCA cycle operation in H. contortus. In a study on the effects of pH and temperature, Kapur et al. (1984), found that SDH exhibits maximum activity (about 120 nmoles/ min/mg protein) at an optimum pH of 8.2 and temperature 32° C. pH optimum for ICDH (about140nmoles/min/mg protein) and MDH (about 30 nmoles/min/mg protein) is 9.0 and 7.8 respectively; optimum temperature being 32º C for both the enzymes (Kapur et al., 1985). The in vitro effects of TMS and RFX on various enzymes at 50 µg/ml, have shown varying degrees of inhibition of SDH and FR. At the same concentration, the activities of other enzymes remained unaltered (Kaur and Sood, 1983c). Also, the in vitro effects of NLZ, ABZ, NLV, NLF, TP and FBZ have been studied on LDH (Kaur and Sood, 1993). Pentose phosphate pathway: This is an alternative pathway of glucose catabolism. There is, however, no definite evidence that this pathway is involved in energy metabolism in parasitic nematodes. Its main function may be to provide NADPH and C5 and C7 sugars for synthetic reactions (Barrett, 1976). In H. contortus, key enzymes of this pathway, viz. glucose- Sood 6-phosphate dehydrogenase (GDPH) and 6phosphogluconate dehydrogenase, as also the in vitro effects of TMS and RFX on these enzymes have been demonstrated (Kaur and Sood, 1985). GDPH exhibits activity (4 nmoles/min/mg protein) at pH 7.4 and 37º C (Kapur et al., 1984). Thermostability of GPDH, separately for male and female worms has been studied (Harpreet et al., 1991). Synthesis of nucleic acids in adult H. contortus, is also indicative of the operation of pentose phosphate pathway (Kapur and Sood, 1986c). Amino acids Amino acids commonly found in proteins, also occur as free acids. There are number of amino acids which are never found as constituents of proteins, but play important metabolic roles. Therefore, it is essential to study both free and bound amino acids. Composition: In adult H. contortus, both free and bound acids are present (Kapur and Sood, 1984b). The level of free amino acids is 191 mg in male and 523 mg in female per 100 g on fresh weight basis. Various acids in free form include aspartic acid (Asp), glutamic acid (Glu), leucine + isoleucine (Leu+Ile), tryosine (Try), lysine+histidine (Lys+His), glycine+serine (Gly+Ser), arginine (Arg), valine+methionine (Val+Met), cystine+cysteine (Cys-Cys+Cys), β-alanine (Ala) and α−Ala in decreasing order of concentration in female, and in male, the order of occurrence is: Asp>Glu>Tyr>βAla>Lys+His>Gly+Ser>Leu+Ile>Val+Met>Agr. Thus, in male Cys-Cys+Cys and α−Ala are missing. In female, concentration of Leu, Ile and Asp is significantly higher than in male, and in male it is of Tyr, β−Ala, Lys+His and Glu. However, concentration of Val, Met, Gly, Ser and Arg is similar in both the sexes. Asp is present in the highest concentration in both the sexes and the acid, in least concentration is Arg in male and α-Ala in female. Like other invertebrates, in H. contortus too, free amino acid pools are dominated by one or two amino acids, i.e., Asp in female and Asp, Glu in male. Proline (Pro) not reported has been earlier demonstrated by Nigam (1979). Higher amount of Met in female than in male indicates the extent of transaminase reactions. More of Lys in male in indicative of its role in maintaining sperm viability (having a histone-like function) as in humans. As in birds, more of Glu in male may play a role in the maintenance of osmolarity and pH of seminal plasma. 9 Haemonchus histochemistry, biochemistry and immunology In both the sexes, bound acids detected are almost similar to those of the free fraction. The differences include the absence of β-Ala and presence of α-Ala, Cys-Cys+Cys, and Pro in both the sexes. Concentration of both total and individual bound acids is far more than that in male. Relative order of occurrence in female is Tyr>Lys+His>Glu>Gly+ Ser>Leu+Ile>Val+Met>Asp>Arg>Cys-Cys+Cys> α-Ala, and in male it isTyr>Gly+Ser>Lys+His>Glu> Leu+Ile> Val+Met>Asp>Cys-Cys+Cys>Arg>α-Ala. However, concentration of Pro is not known. GLUCOSE GLUCOSE -6- PHOSPHATE SERINE TRIOSE PHOSPHATE TRYPTOPHAN PHOSPHOENOL PYRUVATE Metabolism: No significant variation in the composition of amino acids of nematodes inhabiting different environments has been found. This indicates that in nematodes, the composition of the amino acid pool remains largely unaffected, not being dependent only on their absorption form external environment. Thus, the nematodes must be capable of amino acid biosynthesis. This has been demonstrated in many nematodes. Further, in parasitic nematodes, where the major metabolic activities are directed towards egg production, the emphasis on protein and hence amino acid biosynthesis must be considerable. If we are able to find some differences in the enzymes involved in amino acid biosynthesis by the parasite and the host, we can selectively check the synthesis of amino acids in the parasites. Thus, these would not be available for incorporation into egg proteins, and hence the propagation of a species could be checked. Adult H. contortus has been investigated for its ability to utilize various C14-labelled precursors, i.e., glucose, acetate, CO2 and palmitic acid for amino acid biosynthesis (Fig. 2) (Kapur and Sood, 1984c). It has been demonstrated that H. contortus is capable of synthesizing essential as well as non-essential amino acids. Possible mechanisms for the involvement of various precursors in amino acids have been examined. It is not possible to predict whether or not the synthesis of amino acids is at a level commensurate with reproduction. Therefore, further studies need to be carried out on these lines. Also, it would be interesting to elucidate the pathways of amino acid biosynthesis. These studies can give a deeper insight into the metabolism of the worms. ACETATE PHENYL ALANINE TRYOSINE TRYPTOPHAN ALANINE PYRUVATE ACETYL CoA LEUCINE VALINE LEUCINE ISOLEUCINE ALANINE LYSINE ASPARTATE THREONINE SERINE OXALOACETATE GLYCINE CO 2 METHIONINE FUMARATE ARGININE Concentration of α-Ala, Lys+His and Tyr is higher in female and that of Gly, Ser, Cys-Cys+Cys in male. Levels of Leu, Ile, Val, Met, Asp, Glu and Arg are similar in both the sexes. Thus, Tyr is present in the highest concentration and α-Ala the least, in both the sexes. PALMITIC ACID ERYTHROSE-4-PHOSPHATE CITRATE CYSTEINE SERINE CYSTINE TCA CYCLE α - KETOGLUTARATE SUCCINYL CoA ARGININE TRYPTOPHAN GLUTAMATE PROLINE PHENYL ALANNE Fig. 2. Proposed scheme for the involvement of C14-glucose, acetate, carbon dioxide and palmitic acid in amino acid biosynthesis in adults of H. contortus (After Kapur and Sood, 1984; with permission from Elsevier). Absorption/incorporation studies of C14-labelled amino acids in H. contortus by Kocher et al. (2000) have revealed significantly higher absorption of amino acids in females than the males. The absorbed amino acids first get incorporated into cystosolic proteins and after periods of time (30 min), the incorporation in the deoxycholate and sodium dodecyl sulphate extractable membrane bound and nuclear proteins increased significantly. It is postulated that H. contortus absorbs amino acids through cuticle or possibly by ingestion. Although there is no definite proof for the mechanism of ingestion of soluble nutrients, yet it can be similar to the way the parasites ingest blood after attachment to the abomasal mucosa. Anthelmintic effects: The two transaminases- alanine (ALT) and asparate (AST) are involved in transamination, i.e., interconvert a pair of amino acid and a pair of keto acid. ALT is involved in the interconversion of L-alanine to pyruvate, whilst AST converts asparate to oxaloacetate (Rodwell, 1990). Anthelmintic effects of NLZ, TP, NLV, NLF, ALZ and FBZ on these transaminases (Kaur and Sood, 1993), have revealed the increase of these transaminases except with NLZ. Lipids Carbohydrates form the major and possibly the sole energy source of parasitic nematodes. However, the importance of lipids cannot be overlooked, these the 10 being structural and functional constituents. These are important components of membranes, which are in constant stage of dynamic equilibrium. Also, these are incorporated into eggs and are important energy reserves in the free-living stages of animal parasitic nematodes. Thus, it we are able to selectively inhibit lipid biosynthesis in parasites, these would not be available for incorporation into eggs, and we can check the propagation, if not eliminate the parasite. Further, lipid biochemistry (and nutritional requirement) studies indicate the basis of host specificity, which can be exploited for in vitro growth of the parasite. Also, study of lipid biochemistry of different groups of nematodes may reveal phylogenetic relationship or adaptations of parasites. Lipid composition and metabolism in nematodes has been reviewed (Kapur and Sood, 1995) under the titles: composition, absorption/transport, nutritional requirements, biosynthesis, catabolism, lipids in intermediary metabolism, enzymes involved in lipid metabolism and anthelmintic effects. In H. contortus, lipid composition and metabolism has been reviewed (Sood and Kapur, 1989) under the titles: composition, distribution, biosynthesis, catabolism, enzymes involved in lipid metablism, lipids in intermediary metabolism and anthelmintic effects. Composition: Qualitative and quantitative aspects of lipid composition have been studied (Kapur and Sood, 1985). The total lipids constitute 44mg/g of fresh tissue and 200 mg/g of dry tissue. Possibly, higher amount of lipids in H. contortus accounts for the daily production of its own weight of eggs. The non-polar (NP) lipids are present in higher concentration than polar (P) lipids, the NP/P ratio being 1:4. NP lipids include monoacylglycerols (MG), diacylglycerols (DG), sterols, free fatty acids (FFA), hydrocarbons and pigments (Hyd+Pig), triacylglycerols (TG), and sterol esters (S. esters). Of these, TG are present in the highest concentration, followed by FFA, the sterols being the least. Among P lipids, the following components are present in the decreasing order of their occurrence: phosphatidyl choline (PC), phosphatidyl ethanolamine (PE), lyso PE, sphingomyelin+lyso PC, cerebrosides (Cereb.), phosphatidyl inositol (PI) and phosphatidyl serine (PS). Fatty acid analysis of total, P and NP lipids revealed 28, 13 and 15 fatty acids respectively. Fatty acids detected range from C12 to C24, with 1-4 double bonds. C16 and C18 fatty acids constitute more than 50% and C21, C22 about 25% of total fatty acids. An unidentified acid with carbon Sood number 22 or 23 is also present in traces. Metabolism: Nematodes usually contain lipids in considerable amounts, and these are used for energy production, usually under aerobic conditions. It is reasonable to assume that some lipid catabolism could occur in H. contortus because of its aerobic conditions. Adult H. contortus has been investigated for its ability to utilize lipids with regard to total lipids, sterols, FFA, acylglycerols and phospholipids produced during incubation in vitro (Kapur ans Sood, 1987c). All these components exhibit extensive fluctuations, decreasing at some times and increasing at others, thus indicating both biosynthesis and utilization. Also, changes in fatty acid components of total lipids have been analyzed by GLC. Investigations on adult H. contortus for its ability to synthesize various lipids form C14-glycerol (Kapur and Sood, 1987b), have revealed that the worm is capable of utilizing it for lipid biosynthesis. There was significantly more incorporation into P than into NP lipids (P/NP ratio > 1.89). Further the worm is capable of synthesizing all the classes of complex lipids present, viz., acylglycerols, FFA, sterols and S.esters among NP lipids, and PC, PE, PI, PS, sphingomyelin+lyso PC, lyso PE and Cereb among P lipids. Time-course incorporation studies revealed the operation of the MG-pathway for the synthesis of TG. Information has also been obtained indicating which pathways are utilized by the worm for phospholipid synthesis. The results are indicative of de novo synthesis of lyso PC and lyso PE. Incorporation studies with labelled substrates, sodium-1-C14-acetate and U-C14-D-glucose demonstrated that adult H. contortus has extremely active mechainisms for synthesizing all classes of complex lipids including free cholestrol. More of the label from acetate than glucose is incorporated into total lipids. With both the precursors, there is more P than NP synthesis, the P/NP ratio being 3.991 in case of acetate and 1.223 in case of glucose. Among NP components, most of the label from acetate is incorporated into FFA, TG, Hyd+Pig, and label from glucose into TG and FFA only. Among P lipid classes, most of the label from acetate is incorporated into PC and from glucose into PC and PE (Kapur and Sood, 1984a). It is suggested that in order to establish a particular metabolic pathway for lipid biosynthesis, the pool size and turnover lipids must be determined. Haemonchus histochemistry, biochemistry and immunology 11 Accordingly, the presence and functions of the enzymes involved in lipid metabolism must be considered. concentrations greater than 4mm. It was stable at 4ºC for several weeks, but lost 60% of its activity when liated at 60°C for 5 min. Physostigmine and neostigmine inhibited enzyme activity at low (µm) concentrations, whereas phenylmethyl sulfonyl fluoride and sodium fluoride were inhibitory only at high concentrations (Joshi and Singh, 2000). Specific activity of ChE was 33.64± 0.2 (µ moles substrate formed/h/g tissue). After treatment with TMS and RFX, ChE activity was reduced to 31% and 30% respectively (Kaur and Sood, 1982c). H. contortus has also been investigated for its ability to synthesize lipids form simple 14C-precursors (Kapur and Sood, 1986a). It has been shown to have extensively active machanisms for synthesizing all classes of complex lipids present, including free cholestrol from HCO3 and 14C-palmitic acid. With both the precursors, there is more of NP than P lipid synthesis, the NP/P ratio being 1.258 in case of HCO3 and 1.917 in case of palmitic acid. Thus, H. contortus has extremely active mechanisms for the synthesis of complex lipids from exogenously supplied fatty acids from CO2 fixation. Results of the incorporation of carbon from 14Cglycerol into total P and NP lipids (Kapur and Sood, 1991) reveal that radioactivity lost into the medium as lipids decreased during the first 3 h of incubation. This was followed by a drastic increase during the 4th h. Again, there was decline by 75% during the 5th h. However, there was a slight increase in the NP lipids during the 5th h. Enzymes: Lipases hydrolyse esters of long chain fatty acids. In H. contortus, lipases and phospholipases have been detected in adult worms (Kapur and Sood, 1986e). Lipase exhibits an optimum activity at pH 7.4 and temperature of 42° C, and phospholipase at pH 5.0 and temperature of 37° C. An indirect evidence of lipase is also obtained from time-course incorporation studies with glycerol, as decrease in the amount of label in triacylglycerols has been observed (Kapur and Sood, 1987b). Among esterases AChE is involved in nervous transmission in nematodes and choline esterase (ChE) serves to destroy the transmitter, as it does in other groups of animals (Lee, 1965). Both AChE and ChE have been demonstrated in a variety of nematodes, trematodes and cestodes (Barrett. 1981). AChE was purified from extract of adult H. contortus by gel filtration and ion-exchange of ConA-Sepharose chromatography. The enzyme was also secreted by the parasite during in vitro cultivation, and it was partially purified from the excretory-secretory products. The presence of enzyme-specific antibodies was observed in animals infected with H. contortus. The MW of the enzyme by SDS-PAGE was 144 KDa. It showed typical Michaelis-Menten kinetics at low substrate concentrations, but was inhibited by substrate Anthelmintic effects: The in vitro effects of TMS on lipid biosynthesis from simple precursors such as C14acetate and glucose have been studied in adult H. contortus (Kapur and Sood, 1986d). It has a considerable inhibitory effect on lipid biosynthesis, percentage inhibition being 26 in case of acetate and 34 in glucose. Proteins Parasitic helminths are capable of efficient protein synthesis and incorporation of labelled amino acids into proteins has been demonstrated in several species. Adult H. contortius is capable of efficient protein synthesis from acetate, glucose, palmitic acid and CO2 (Kapur and Sood, 1986c). Of these, palmitic acid is the most efficient precursor and CO2, the least. Extent of synthesis from acetate and glucose is intermediate. Nucleic acids Adult H. contortus is capable of efficient nucleic acid biosynthesis from acetate, glucose, palmitic acid and CO2 (Kapur and Sood, 1986d). Synthesis from acetate and CO2 is negligible, compared to that from glucose and palmitic acid. It is suggested that glucose is partially decomposed via HMP-pathway into ribose and hence incorporated into RNA. Nutrition Digestive enzymes: The nutritive functioning of H. contortus depends on the activity of the digestive enzymes, especially protease (s), which play a number of critical roles such as digestion of host proteins, retardation of blood coagulation, evasion of immune response, invasion of host tissues and blood feeding and its digestion (see Kocher et al., 2002a). In a study on the characterization and purification of cystosolic and membrane-bound protease (s) in adult H. contortus, different fractions showed optimum protease activity at 37° C, pH 8.5 and 8.0 mg casein of 12 concentration (Kocher et al., 2002a). The female fractions had a particularly high activity of protease (s) in comparision with the male fractions, especially of membrane-bound enzymes in the anterior half. Inhibition/activation studies revealed the presence of four kinds of protease (s) in the cystosolic and membrane-bound fractions. Protease (s) in different fractions are purified to a greater extent by higher concentrations of saturated ammonium sulphate solutions, i.e., ranging from 50 to 65%. The purification study revealed the presence of multiple forms of protease (s) in cystosolic and membranebound extracts of H. contortus. Studies have also been made to elucidate the inhibition of protease (and lipase activity) from excretorysecretory products, cystosolic and membrane bound fractions of male and female H. contortus by immunoglobulins raised in rabbits against cytosolic fraction (Kocher et al.,1996). Biochemistry in taxonomy Possibility of the use of electrophoresis in polyacrylamide gel for taxonomy of helminths is well established. Electrophoretic analysis of proteins of knobbed and linguiform morphs of female H. contortus revealed marked differences (Sood and Kapur, 1982a). Proteins of knobbed form have 13 fractions, and those of linguiform 14. Further, in knobbed form, there are 3 bands at cathode and 2 at anode, while in linguiform morph, at cathode, there are 5 bands and at anode only one. This observation (along with other evidences) tends to support the view of subspeciation in H. contortus. Miscellaneous Glutamate dehydrogenase (GDH) plays a central role in amino acid deamination and in the formation of αamino nitrogen groups form ammonia. Its optimum activity (162 nmoles/min/mg protein) is at pH 7.0 and temperature of 27 ° C (Kapur et al., 1984). Thermostability of GDH has also been studied (Harpreet et al., 1991. The in vitro effects of NLZ, ABZ, NLV, NLF, TP and FBZ have been studied in AkPase and AcPase. All the six drugs caused decrease in the enzyme levels of AkPase, the maximum effect being exerted by NLZ. In AcPase, marked decrease in the level was observed, maximum effect being caused by NLF (Kaur and Sood, 1993). Malic enzyme (ME) is one of the enzymes involved in CO2 fixation and its presence suggests an alternate fate Sood of phosphoenolpyruvate and ATP is generated, when the latter is reduced to pyruvate. Activity of ME has been studied in adult H. contortus (Kaur and Sood, 1982b). The in vitro treatments with TMS had no significant effect. However, after RFX treatment, ME activity was reduced to 38%. Kaur and Sood (1982a) studied the effect of TMS and RFX on total volatile fatty acids under in vitro conditions. IMMUNOLOGY Chemotherapy is an inadequate means of controlling haemonchosis because of the development of resistance to anthelmintics. In view of the severe pathogenicity and economic losses due to haemonchosis, various other strategies, including breed resistance, biological control and immunological control through vaccine are being studied. Immunology is an exciting area for formulating new products for therapeutic and diagnostic purposes. In immunological control methods, with vaccines as targets, specific immunodominant antigens play a significant role. Immunoparasitology of Haemonchus infestation has been reviewed (Kapur et al., 2001). The topics discussed are: vaccination, genetic control, immune responese, transfer of immunity, immunodiagnostics and host effects. The various aspects of H. contortus immunology, reported from India are as follows: Kaur et al. (2002a) used Dot-ELISA to study kinetics of hyperimmune sera of rabbits immunized with adult H. contortus. A high titre sera were obtained in immunized rabbits and observed to be maintained for 180 days, in response to adult H. contortus soluble extract which can be of significant value in the identification of immunodominant antigens and their further characterization using natural host sera. Earlier studies of Sood et al. (1996) have shown that competitive-inhibition Dot-ELISA using adult H. contortus antigen is able to detect as low as 10 pg of the antigen and can be suitably applied at the field leval for mass screening of infected animals. To enhance this sensitivity Kapur-Ghai et al. (2004) have developed avidin-biotin ELISA for detection of H. contortus antigens. Immunodominant antigens of adult H. contortus that can evoke a protective immune response in the host have been identified (Kaur et al., 2002b). Since the Haemonchus histochemistry, biochemistry and immunology 13 antigenic component with MW 91.2 KDa was recognized in the immune sera of all the rabbits and also in sera collected at intervals, it is suggested as the immunodominant component of adult H. contortus. and smooth gave three precipitin lines with rabbit antisera. Also, lines unite in a manner indicating marked antigenic differences between the three types. Since serological techniques are considered to be sensitive tools in taxonomy, the present work has indicated that taxonomic importance should be attached to the vulvar configurations in female H. contortus. Two low molecular weight proteins of 15 and 22 KDa were isolated from the extract of adult H. contortus by gel filtration conA-sepharose and affinity chromatography on antibody-sepharose. These proteins were also indentified in the excretorysecretory products of adult parasites. Antibodies against these proteins were identified in the sera of animals infected with H. contortus. Upon immunization, these proteins protected animals against challenge H. contortus infection, as egg shedding and worm burden significantly reduced. The importance of these antigens in the host-parasite relationship has been discussed (Joshi and Singh, 1999). Preliminary studies carried out by Kocher et al. (2002b) to determine the relationship between level of circulating immune complex (CIC) and the possibility of ascerting mortality caused by H. contortus infestation in sheep and goat, have revealed that determination of CIC level may prove to be important parameter for early diagnosis of the state of the host immunized with H. contortus. To detect anthelmintic resistance using thiabendazole, a larval development assay (LDA) has been standardized. A linear dose-response relationship was observed between the probit of larval development of the logarithms of anthelmintic concentration of both egg hatch assay and LDA (Singh et al., 2003). Sood and Kapur (1981) observed immunological changes in the spleen of rabbits in response to antigens of H. contortus female. The study of sections showed hyperplasia of reticuloendothelial cells and duplication of red pulp. Haemosidrin pigment was also observed. A few eosinophils and neutrophils were also seen at the cortical region. Antibodies being proteins, their synthesis is similar to that of other proteins and hyperplasia of reticuloendothelial cells indicates increased protein synthetic activity. In support of the biochemical techniques in taxonomy in H. contortus (Sood and Kapur, 1982b), immunodiffusion patterns of antigens from phenotypically different females have been studied (Sood and Kapur, 1982c). The studies indicate that the three phenotypes---smooth, linguiform, and knobbed differ from each other serologically. Linguiform antigens gave five precipitin lines, knobbed four lines REFERENCES Anonymous. 1992. Distribution and impact of helminth diseases of livestock in developing countries. FAO animal production and health paper 96. Food and Agriculture Organization of the United Nations, Rome. Barrett B. 1976. Bioenergetics in helminths. In: Biochemistry of Parasites and Host-Parasite Relationship. H van Den Bossche (Edt.). North Holland Biomedical Press, Amsterdam. Barrett J. 1981. Biochemistry of Parasitic Helminths. MacMillan Publisher, Ltd., London and Basingstoke. Bird AF. 1971. The Structure of Nematodes. Academic Press, London. Chitwood BG and Chitwood MB. 1974. Introduction to Nematology, University Press, Baltimore, Maryland. Chopra AK and Premvati G. 1977. Glucose metabolism and lactic acid production in sheep nematodes. Indian J Parasita 1:93-96. Fraser CM. 1991. The Merck Veterinary Manual. A handbook of diagnosis, therapy and disease prevention and control for the veterinarians. Merck and Co. Inc. Rahway, USA, pp.205-215. Harpreet, Sood ML and Soni GL. 1991. Thermostability of some enzymes of Haemonchus contortus and Trichuris trichura. Indian J Parasit 15: 171-173. Johl M. 2003. Histomorphological variations in some gastrointestinal nematodes of domestic ruminants. In: Helminthology in India. ML Sood (Edt.). International Book Distributors, Dehra Dun , India, pp.453-466. Joshi P and Singh BP. 1999. Isolation and characterization of two low molecular weight protective antigens of Haemonchus contortus. Indian J Anim Sci 69: 284-288. Joshi P and Singh BP. 2000. Purification and characterization of cholinesterase from Haemonchus contortus. Indian J Biochem Biophys 37: 192-197. Kapur J, Parmar A and Sood ML. 2001. Immunoparasitology of Haemonchus infestation. Prof. VN Capoor Comm.Vol. Sandeep Kumar Malhotra (Edt.). Ankit Publication, Allahabad, pp. 26-34. Kapur J and Sood ML. 1984a. Haemonchus contortus: Lipid biosynthesis from C14-labelled acetate and glucose. Zbl Vet Med B 31:225-230. 14 Sood Kapur J and Sood ML. 1984b. Amino acid composition of the adults of Haemonchus contortus (Nematoda: Trichostrongylidae). Helminthologia 21:267-273. Kapur J and Sood ML. 1984c. Amino acid biosynthesis in Haemonchus contortus from C14-labelled precursors, in vitro. Vet Parasitol 15:193-199. Kapur J and Sood ML. 1984d Effect of pH and temperature on four dehydrogenase enzymes of Haemonchus contortus. Acta Vet Hung 32:165-170. Kapur J and Sood ML. 1985. Haemonchus contortus: Qualitative and quantitative analysis of lipids. Zbl Vet Med B 32 :345-353. Kapur J and Sood ML. 1986a. Haemonchus contortus: Lipid biosynthesis from C14 labelled palmitic acid and sodium bicarbonate. Acta Vet Hung 34:73-80. Kapur J and Sood ML. 1986b. Metabolism of C14-labelled glucose, acetate and palmitic acid to CO2 in adult Haemonchus contortus (Nematoda : Trichostrongylidae). Indian J Parasit 10:85-86. 14 Kapur J and Sood ML. 1986c. Incorporation of carbon from C labelled precursors into major chemical fractions of Haemonchus contortus, in vitro. Acta Vet Hung 34:183-187. Kapur J and Sood ML. 1986d. Effect of dl-tetramisole on lipid biosynthesis in adults of Haemonchus contortus (Nematoda : Trichostrongylidae). India J Comp Microbiol Immunol Infect Dis 7:10-13. Kapur J and Sood ML. 1986e. Effect of pH and temperature on lipase and phospholipase of adult Haemonchus contortus (Nematoda : Trichostrongylidae). J Parasit 72:346-347. Development of avidin-biotin based ELISA for detection of Haemonclus contortus (Nematoda: Trichostrongylidae) antigens. Indian J Comp Microbiol Immunol Infect DIS 25:45-46. Kaur K, Kapur J, Parmar A and Sood ML. 2002a. Kinetics of antibody response by Dot-ELISA in rabbits immunized with adult Haemonchus contortus antigen. Parasite 9:363-365. Kaur K, Kapur J, Parmar A and Sood ML. 2002b. Identification of immunodominant antigens of adult Haemonchus contortus (Nematoda : Trichostrongylidae). J Vet Med 49:260-262. Kaur M and Sood ML. 1990. Effect of Nilzan® and albendazole on the absorptive surfaces of Haemonchus contortus (Nematoda) -a histoenzymic study. Angew Parasitol 31:167-181 Kaur M and Sood ML. 1992a. In vitro effect of Nilverm and Nilverm forte on the absorptive surfaces of Heamonchus c o n t o r t u s ( N e m a t o d a : Tr i c h o s t r o n g y l i d a e ) . Helminthologia 29:31-37. Kaur M and Sood ML. 1992b. Histoenzymic effects of thiophenate and fenbendzaole on the absorptive surfaces of Haemonchus contortus. Acta Vet Hung 40:63-70. Kaur M and Sood ML. 1993. In vitro anthelmintic effects on some enzymes of Haemonchus contortus and Trichuris globulosa. Prof. SP Gupta Comm. Vol. Indian J Helminth 45:26-35. Kaur M and Sood ML. 1996. In vitro effects of anthelmintics on the histochemistry of Haemonchus contortus and Trichuris globulosa. Appl Parasitol 37: 302-311. Kapur J and Sood ML. 1987a. Biochemistry of Haemonchus- a review. Angew Parasitol 28:211-228. Kaur R and Sood ML. 1982a. Haemonchus contortus: The in vitro effects of anthelmintics on total glucose and glycogen contents and total volatile fatty acids. Acta Vet Hung 30:265-269. Kapur J and Sood ML. 1987b. Lipid biosynthesis from C14glycerol in Haemonchus contortus (Nematoda : Trichostrongylidae), in vitro. Acta Vet Hung 35:259-265. Kaur R and Sood ML. 1982b. Haemonchus contortus. The in vitro effects of dl-tetramisole and rafaxonide on glycolytic enzymes. Int J Parasit 12:585-588. Kapur J and Sood ML. 1987c. Changes in lipids and free fatty acid fraction in adult Haemonchus contortus during incubation in vitro. Vet Parasitol 23:95-103. Kaur R and Sood ML. 1982c. In vitro effect of anthelmintics on malic enzyme and cholinesterase of Haemonchus contortus (Nematoda : Trichostrongylidae). Indian J Parasit 6:267268. Kapur J and Sood ML. 1991. Research Note. Secretion and excretion of endogeneously synthesized lipids by adlut Haemonchus contortus in vitro. J Helminth 65:73-75. Kaur R and Sood ML. 1982d. In vitro effect of anthelmintics on the phosphatases of Haemonchus contortus (Rud., 1803). Indian J Parasit 6: 279-274. Kapur J and Sood ML. 1995. Lipid composition and metabolism in nematodes. In : perspectives in Nematode Physiology & Biochemistry. J Kapur and ML Sood (Edts.). Narendra Publishing House, Delhi, India, pp. 97-154. Kaur R and Sood ML. 1983a. Effects of anthelmintics on the absorptive surf aces of adult Haemonchus contortus in vitro: a histoligical study. Folia Parasit 30:146. Kapur J and Sood ML and Soni Gl. 1984. Effect of pH and temperature on four dehydrogenase enzymes of Haemonchus contortus. Acta Vet Hung 32:165-170. Kaur R and Sood ML. 1983b. Review article. Carbohydrate metabolism in Haemonchus contortus (Nematoda : Trichostrongylidae). Indian J Parasit 7:125-132. Kapur J, Sood ML and Soni Gl. 1985. Haemonchus contortus: Effect of pH and temperature on some dehydrogenases. Acta Vet Hung 33:19-23. Kaur R and Sood ML. 1983c. The effects of dl-tetramisole and rafoxanide on tricarboxylic acid cycle enzymes of Haemonchus contortus in vitro. Vet Parasitol 13:333-340. Kapur-Ghai J, Thapar V, Kaur K and Sood ML. 2004. Kaur R and Sood ML. 1985. In vitro effect of anthelmintics on Haemonchus histochemistry, biochemistry and immunology 15 pentose phosphate pathway enzymes of Haemonchus contortus (Nematoda :Trichostronogylidae). Indian J Parasit 9:159160. Sood ML and Kapur J. 1980. Research note. Inorganic elements in the adults of Haemonchus contortus (Nematoda : Trichostrongylidae).J Helminth 54:253-254. Kaur R and Sood ML. 1986. Effects of anthelmintics on Haemonchus contortus (Nematoda : Trichostrongylidae). Vet Res Comm 10:21-36. Sood ML and Kapur J. 1981. Immunologic changes in the spleen of rabbits in response to antigens of Haemonchus contortus female (Nematoda : Trichostrongylidae). Folia Parasit 28:272. Kocher DK, Ahuja SP and Sood ML. 1996. Proteases and lipases as host protective antigens in Haemonchus contortus. In:Parasitic Diseases-new horizons. Proc Eighth Nat Cong Vet Parasit Nat Symp Mol Parasit. DP Banerjee, JD Ghosh and SK Gupta (Edts.). Department of Veterinary Parasitology, CCS Haryana Agricultural University, Hisar pp. 82-86. Kocher DK, Ahuja SP and Sood ML. 2000. Absorption and incorporation of 14C-labelled amino acids in Haemonchus contortus. J Vet. Parasit 14 : 129-131. Kocher DK, Ahuja SP and Sood ML. 2002a. Charaterization and purification of cystosolic and membrane-bound protease (s) in adult s of Haemonchus contortus. J Vet Med B 49:71-75. Kocher DK, Ahuja SP and Sood ML. 2002b. Relationship between circulating immune complex (CIC) level and mortality of host immunized with Haemonchus contortus. J Parasit Dis 26:46-47. Lee DL. 1965. The Physiology of Nematodes. Oliver and Boyd, Edinburgh and London. Sood ML and Kapur J. 1982a. Haemonchus of ruminants- a bibliography of titles prepared from Helminthological Abstracts covering the period 1932-1980. Shikha Publications, Ludhiana, India. Sood ML and Kapur J. 1982b. Electrophoretic analysis of proteins of knobbed and linguiform morphs of female Haemonchus contortus (Nematoda : Trichostrongylidae). Helminthologia 19:273-278. Sood ML and Kapur J. 1982c. Haemonchus contortus: Immunodiffusion patterns of antigens from phenotypically different females. Expl Parasit 53:164-169. Sood ML and Kapur J. 1989. Review article. Lipid composition and metabolism of Haemonchus contortus (Nematoda :Trichostrongylidae). Indian J Parasit 13:367371. Sood ML and Kaur C. 1976. Studies on vulvar configurations in Haemonchus contortus (Rud., 1803) from goats at Ludhiana, India. Riv di Parassit 37:13-33. Nigam SC. 1979. Amino acid composition of nematode parasites. Indian J Helminth 31:69-71. Sood ML and Kaur C. 1977. Morphological and histological studies on the vulvar configurations in Haemonchus contortus (Rud., 1803). Folia Parasit 24:111-115. Premvati G and Chopra AK. 1979. In vitro variation of glycogen content in three sheep nematodes. Parasitology 78:355-359. Sood ML, Kaur G, Parmar A and Kapur J. 1996. Development of Dot-ELISA for detection of Haemonchus contortus antigen. Helminthologia 33:73-75. Rodwell VW. 1990. Harper's Biochemistry. RK Mdrray, DK Ranner, PA Mayes and YW Rodwell (Edts.). 22nd ed., Prentice Hall International Inc. Connecticut. Sood ML and Kaur M. 1983. Morphology and histochemistry of the spicules and gubernaculum of Haemonchus contortus (Nematoda : Trichostrongylidae). Folia Parasit 30:249-255. Singh D, Swarnkar CP, Khan FA and Bhagwan PSK. 2003. Standardization of larval development assay for detection of anthelmintic resistance in Haemonchus contortus. J Vet Parasit 17:21-25. Sood ML and Kaur R. 1982. The in vitro effects of some drugs on the morphology and histochemistry of adult Haemonchus contortus (Nematoda : Trichostrongylidae). Helmin-thologia 19:61-70. Singh J and Johl M. 2001. Observations on the foregut (stomodaeum) of Haemonchus contortus Rud., 1803. Uttar Pradesh J Zool 21:139-145. Sood ML and Sehajpal K. 1978. Morphological, histochemical and biochemical studies on the gut of Haemonclus contortus (Rud., 1803). Z Parasitenkd 56 : 267-273. Sood ML. 1981. Haemonchus in India. Parasitology 83: 639650. Sood ML, Sood M and Sood ML. 1999. Experimental determination of diffusion coefficients of NaCl and KCl in Haemonchus contortus at 298.16K. J Indian Chem Soc 76:410-411. Sood ML. 1999. Absorptive surfaces of Haemonchus contortus (Nematoda : Trichostrongyloidea). JPAS 1 : 225-227. Sood ML. 2003. Haemonchosis in India. In: Helminthology in India. ML Sood (Edt.). International Book Distributors, Dehra Dun, India, pp. 575-617. Sood ML and Kalra S. 1977. Histochemical studies on the body wall of nematodes : Haemonchus contortus (Rud., 1803) and Xiphinema insigne Loos, 1949. Z Parasitenk51 : 265273. Ward PFV. 1982. Aspects of helminth carbohydrate metabolism. Parasitology 84:177-194. Wright KA. 1987. The nematodes' cuticle---its function, homology, analogy-a current consensus. J Parasit 73:10771083. Review Journal of Parasitic Diseases: June 2006, Vol. 30, No. 1, 16–29 J PD Immunological perspectives and malaria vaccine H. S. Banyal and N. Elangbam Department of Biosciences, Himachal Pradesh University, Shimla. ABSTRACT. Even after more than a century of efforts to either eradicate or control malaria, it still remains a major vector borne parasitic disease that affects nearly a third of world's population, mainly in poor developing countries. Increasing incidences of parasite resistance to available antimalarial drugs and of mosquitoes to commonly used insecticides necessitates an alternative combat strategy for the control of malaria. The development of an effective malaria vaccine is one such strategy. Herein, we focus on various immunological aspects of malaria including target antigens that can serve as potential candidate vaccines, and on the status and prospects of the development of a malaria vaccine. Keywords: immune response, malaria, Plasmodium, target antigens, vaccine INTRODUCTION Malaria, derived from the Italian word for bad air was attributed to fevers in populations living in the vicinity of marshes. Edwin Smith Surgical Papyrus 1600 B. C., indicated about the disease while the Greek physician, Hippocrates gave the first accurate clinical description of malarial fever in 400 B. C. (Boyd, 1949). Malarial fevers were known in ancient China and Arabian countries, and have also been mentioned in ancient Indian literature like 'Charaka Samhita'. Meckel in 1847 was the first person to observe black granules embedded in protoplasmic masses in the blood of a severely ill malarious patient. In 1880, a French army physician, Charles-Louis-Alphonse Laveran observed exflagellation of a parasite and described it as Laverania falcipara in 1884. Manson suggested mosquitoes to be the host for malaria parasite's extrinsic development which encouraged Sir Ronald Ross to investigate the fate of malaria parasite in various mosquito species in India. In 1898, Corresponding author: Prof. H. S. Banyal, Laboratory of Parasitology and Immunology, Department of Biosciences, Himachal Pradesh University, Shimla-171 005, H. P., India. E-mail: hsbanyal@yahoo.co.in Ross succeeded in completely elucidating the sporogony of Plasmodium relictum in Culex pipens fatigans. Meanwhile, Italian worker Bignami in 1899 succeeded in infecting a healthy volunteer with Plasmodium falciparum through the bites of mosquito. At the same time, sporogony in P. falciparum and P. vivax was elucidated in anopheline mosquitoes (Bastianelli and Bignami, 1899; Grassi et al., 1899a), and also the development of sporogonic stages of P. malariae in Anopheles claviger (Grassi et al., 1899b). Differential descriptions of P. vivax and P. malariae were given by Grassi and Feletti (1892) and for P. falciparum by Welch (1897). The fourth human malaria parasite species, P. ovale was described in 1922 (Stephens, 1922). Even after more than a century of efforts to either eradicate or control malaria, it still remains a major global health hazard and one of the most important vector borne human diseases prevalent in more than107 countries that affects nearly a third of the world's population. Malaria is prevalent mainly in poor developing countries and causes more deaths than any other parasitic disease with sub-Saharan Africa accounting for nearly 90% of the cases. Other places like South-East Asia, Oceania, Middle East and Malaria vaccine development Latin America also face serious malaria problems. In Africa, every 30 seconds, a child dies of malaria. Various strategies have been adopted to control the spread of malaria through vector elimination and chemotherapy. Taking into account the increasing resistance of Plasmodium to chemotherapeutic agents, and of Anopheles to conventional insecticides, there is a critical need for an effective malaria vaccine to combat malaria. Great advances have been made in understanding the immunological perspectives of malaria which should help in the fight against the disease. LIFE CYCLE OF MALARIA PARASITE Malaria is caused by a protozoan parasite, Plasmodium, four species of which infect humans: Plasmodium falciparum, P. vivax, P. ovale and P. malariae. Mortality and morbidity are mainly due to P. falciparum, although P. vivax is more widespread geographically. Malaria parasite has a complex life cycle alternating between a vertebrate, ranging from a reptiles to mammals, and an arthropod host female anopheline mosquitoe. Each bite of an infected female Anopheles inoculates 5-20 sporozoites which, within 30 min, find their way into hepatocytes. Each sporozoite multiplies and differentiates intracellularly into a liver stage trophozoite and ultimately a schizont. Rupture of hepatocytes releases merozoites (Mzs) into circulation, which continue a cycle of red cell invasion and multiplication causing clinical manifestations of the disease. Within erythrocyte, each Mz develops into a trophozoite that matures and divides, generating a schizont that gives rise to up to 32 Mzs within 48 or 72 h depending upon the species. The Mzs upon invading new erythrocytes either maintain the blood schizogony or some of them differentiate into male or female gametocytes which are ingested by the blood feeding mosquito. In the mosquito gut, the gametocytes emerge as gametes and fertilize to produce motile ookinetes which burrow into the gut wall of mosquito to form oocysts. Finally, sporozoites released into the body cavity ultimately find their way to the salivary glands, and are injected to a new host during the next blood-meal. IMMUNOLOGY OF MALARIA Malaria infection gives rise to immune responses by the host which are regulated both by the innate and adaptive immune systems. Immunity to malaria involves both cell-mediated and humoral immune mechanisms through both T-cells and B-cells. The 17 cellular and humoral arms of immunity are tightly bound through cytokines which control the immune response with both antibody and cellular immunity playing critical roles in protective immunity. The mechanisms of non-specific innate defense are poorly defined. Neutrophils, mononuclear phagocytes and natural killer (NK) cells appear to play an important role(s) in innate immunity to malaria infections. Humoral immunity: B-cells are primarily concerned with production of antibodies, which form humoral immune response. In malaria endemic areas, induction of strong humoral immune responses involving predominantly IgM and IgG have been reported (Chelimo et al., 2005; Couper et al., 2005). Antibodies act against different stages of parasites, and are predominantly effective against erythrocytic stages. IgG 1 and IgG 3, the two cytophilic isotypes in humans, predominate in malaria protected individuals and IgG2a in P. yoelii has been reported to protect rodents (Druilhe et al., 2005). The possible mechanisms of action of antibodies include inhibition of Mz invasion of erythrocytes and inhibition of intraerythrocytic development of parasites or both. Antibodies can also cause neutralization and agglutination of Mzs. Protective antibodies have very limited direct effect on parasite growth and invasion but act in co-operation with blood monocytes, known as antibody-dependent cellular inhibition (ADCI). In ADCI, soluble mediators are released at the time of schizont rupture triggered by the contact between Mz surface components and cytophilic antibodies bound to monocytes which diffuse in the serum and block multiplication of surrounding parasites at the uninucleate stage (Bouharoun-Tayoun et al., 1995). Tumor necrosis factor-α (TNF-α) has been implicated in ADCI and is thought to inhibit the ring stages of parasite development. Cell-mediated immunity: The importance of cellmediated response in parasitaemia suppression in animal models was suggested by the resolution of acute malaria infection by several Plasmodium sp. in B-cell deficient but not athymic mice (Grun and Weidanz, 1981; Cavacini et al., 1990). Acquisition and maintenance of protective immunity to malaria is T-cell dependent as T-cells are essential both in regulating antibody formation and in inducing antibody independent immunity (Webster et al., 2005). T-cells act as helpers for antibody response but also as effector cells as they can inhibit parasite growth 18 in vitro. The cell-mediated immune effector mechanisms include macrophage activation by interferon-γ (IFN-γ ) derived from γ T cells, NK cells or T helper1 (Th 1) and inhibition of parasite growth and development inside hepatocytes mediated by CD8+ cytotoxic and IFN-γ producing T cells (Tsuji and Zavala, 2003). Cell-mediated immune responses may protect against both pre-erythrocytic and erythrocytic stage parasites. Immunity against pre-erythrocytic stages: In preerythrocytic stage immunity, infected hepatocytes expressing major histocompatibility complex (MHC) molecules are the primary target of cell-mediated immune responses. Both CD 4+ T-cells and CD8+ Tcells recognize parasite derived peptides presented by class I and II MHC molecules, respectively, present on the surface of infected hepatocytes. CD8+ T-cells are thought to be the primary mediators against preerythrocytic stages (Meraldi et al., 2005). It has been suggested that CD 8+ T-cells interact with MHCpeptide complex on the surface of infected hepatocytes and secrete IFN-γ, which in turn induces the infected hepatocytes to produce nitric oxide (NO) that renders the parasite non-infectious. CD8+ T-cells rather than CD8+ cytotoxic T lymphocytes (CTLs) per se, are the critical effector cells of pre-erythrocytic stage immunity. Immunity against erythrocytic stages: Erythrocytic stage immunity is thought to involve both antibodies and T-cell. Antibodies are presumed to work against parasite proteins on the surface of erythrocytes and prevent the sequestration of parasites in microcirculation resulting in their destruction by spleen. Anti-Mz antibodies may also act in other protective mechanisms like complement-mediated lysis and act through co-operation with Fc bearing cells. Parasite proteins present intracellularly or expressed on the surface of Mzs are also target of antibodies that prevent invasion of erythrocytes by parasites (Banyal and Inselburg, 1985). In humans, cytophilic antibodies IgG1 and IgG3 may bind to Mzs and thus facilitate their uptake by phagocytes or mediate antibody-dependent cellular cytotoxicity (ADCC) or ADCI (Tebo et al., 2001). Pouvelle et al. (1991) reported that antibodies can penetrate the infected RBC through the parasitophorous ducts and bind to the intracellular parasite. Studies in animal models have implicated T-cell mediated, antibody-independent mechanism(s) in Banyal and Elangbam immunity against blood-stage malaria parasites. CD4+ T-cells are essential for immune protection against asexual blood stages in both murine and human malaria infections. Adoptive transfer of CD4+ T-cells from immune donor mice to nude mice suppresses acute infection. The antibody-independent mechanisms of action of CD4+ T-cells have been suggested to involve cytokines and NO and T-cells (Seixas et al., 2002). In P. chabaudi infection T-cells are stimulated during primary infection and their expansion is observed during recovery from acute infection. CD4+ T-cells act through the release of cytokines that may exert parasiticidal or parasitostatic effect, activate macrophages and provide help for antibody production by B-cells. Activated T-cells but not T-cells from malaria naïve donors inhibit parasite replication in erythrocytes in vitro, suggesting their protective function (Perlmann and Troye-Blomberg, 2002). In humans, T-cells act not only as helpers for an antibody response but also act as effector cells as they can inhibit parasite growth in vitro. As human erythrocytes do not express MHC antigens, lysis of infected-erythrocytes by CD8+ cytotoxic T lymphocytes (CTLs) plays no role in protection against blood-stage parasites. Sterile immunity induced by repeated low grade infection of red cells without any detectable antibodies suggests that T-cell mediated protection operates in humans (Pombo et al., 2002). Role(s) of cytokines: In malaria, parasite killing requires the production of inflammatory cytokines like IFN-γ, interleukin (IL)-1 and IL-6 and others that can have deleterious systemic effects and have been correlated with malaria pathology. The cytokines TNF-α, IL-1 and IL-6 have been connected with severe malaria (Artavanis-Tsakonas et al., 2003). In murine malaria infection, TNF-α and IFN-γ activate macrophages to phagocytose parasitized-erythrocytes and release NO that causes destruction of parasite which results in protection from severe disease (Good and Doolan, 1999). In humans, TNF-α, IFN-γ and NO are associated with the resolution of fever and parasite clearance (Kremsner et al., 1996). However, high levels of circulating TNF-α are associated with severe P. falciparum malaria and high levels of IFN-γ with fever. IL-12, produced by mononuclear phagocytes is involved in protection against pre-erythrocytic and blood-stage infection by a Th1 anti-malaria response (Doolan and Hoffman, 1999). In contrast, anti- 19 Malaria vaccine development inflammatory cytokines like IL-10 and transforming growth factor-β (TGF-β) counteract the production and cytopathic effects of the pro-inflammatory cytokines (Omer et al., 2000). Lower than normal levels of circulating TGF-β are seen in patients with symptomatic P. falciparum infection. Also risk of febrile illness is associated with high ratios of IFN-γ, TNF-α or IL-12 to TGF-β (Dodoo et al., 2002) which emphasize the importance of the balance between proand anti-inflammatory cytokines. TGF-β plays an essential role in down regulating the production of potentially pathogenic pro-inflammatory cytokines and may be a novel mechanism of pathogen- mediated TGF-β activation. TNF plays a central role both in protection and pathogenesis of malaria, whereas elevated levels of serum IFN-γ enhance production of TNF, NK and T-cells and have been identified as potential sources of IFN-γ in malaria infection. NO can modulate anti-microbial activity, smooth muscle contraction, cytokine production and has been correlated with malaria immunity as well as pathogenesis (MacMicking et al., 1997). NO has also been implied in protection from blood-stage malarial parasites in humans as plasma NO levels increased in patients with P. falciparum and P. vivax. Target antigens: Problems of the resistance to chemotherapeutic agents and insectides emphasize the urgency of a suitable malaria vaccine. The complex life cycle of malaria parasite with distinct developmental stages expressing multiple antigens could provide targets of immune responses. There are essentially six targets for a malaria vaccine in the whole life cycle of the parasite namely, sporozoites, liver stages, Mzs, infected-erythrocytes, parasite toxins and sexual stages. Sporozoite: In 1967, Nussenzweig and co-workers showed that mice immunized with radiation attenuated sporozoites of P. berghei were protected against challenge with infectious sporozoites. This immunization confers sterile protective immunity which is species-specific but not strain specific (Nussenzweig et al., 1967). Immunization with heat killed, formalin treated or lysed sporozoites could not induce protection emphasizing the requirement for live sporozoites in the hepatocytes for protective immunity. Intrahepatocytic parasites are a major target of protective immunity induced by immunization with irradiated sporozoites and CD8+ T-cells specific for epitopes of parasite proteins expressed in the hepatocytes are considered primary effectors with IFN-γ playing a major role (Plebanski and Hill, 2000). Circumsporozoite proteins: Protective antibodies against sporozoites are mainly directed against circumsporozoite proteins (CSP). A 44K CSP exhibited protection against P. berghei (Potocnjack et al., 1980). Passive protection by polyclonal antibodies against central repeat has also been reported for P. berghei and P. yoelii (Egan et al., 1987; Wang et al., 1995). P. vivax CS-derived synthetic peptides have shown good antigenicity and immunogenicity eliciting both humoral and cellular responses (Herrera et al., 2004; 2005). Liver stage antigens (LSA): Immunity during this stage is mostly mediated by cellular-dependent mechanisms involving CD8+ T-cells, CD4+ T-cells, NK cells and T-cells. Studies in mice indicate that IFN-γ produced by activated CD8+ T-cells induce infected- hepatocytes to synthesize NO which has potent anti-parasitic activity (Doolan and Hoffman, 2000). Some of the identified liver stage antigens include LSA-1, LSA-2 and LSA-3. LSA-1 is expressed specifically in liver stage parasites and no homologue has been identified in mouse or nonhuman primate malarias. It is a 200 kDa protein with conserved sequence across strains and involved in naturally acquired responses in human protection. LSA-1 specific protective immune responses includes CD8+ T-cells, IFN-γ, IL-10 and antibodies in natural transmission. Daubersies et al. (2000) reported protection of chimpanzees against P. falciparum by immunization with LSA-3, 200 kDa protein expressed both in sporozoites and liver stages and is highly conserved. Pf LSA-3 DNA immunization induced potent Th 1 response with protective properties and conferred protection against P. yoelii challenge in mice. Sporozoite surface protein 2 (SSP-2): SSP-2 is also known as thrombospondin-related anonymous protein (TRAP) involved in sporozoite invasion and is carried into hepatocytes along with CSP ( Muller et al., 1993). SSP-2 contains a sulphated glycoconjugated binding peptide sequence needed for parasite invasion. Antibodies to SSP-2 prevent sporozoites from invading hepatocytes in vitro. Merozoite The Mz is the only extracellular stage of the malaria 20 parasite in the human host other than the sporozoite making it a visible target for antibodies. Antibodies targeting Mz proteins, mainly its surface proteins, interfere in the invasion process through agglutination followed by phagocytosis or blocking of Mzerythrocyte interaction. Several Mz surface proteins have been identified and some are targets for leading malaria candidate vaccines. Mz surface proteins (MSP): MSP-1 is the most abundant protein on the surface of Mzs (Polley et al., 2003). It is a 190230 kDa protein on the surface of Mz that is processed into smaller fragments at the time of invasion of erythrocytes and exists as a non-covalently linked complex of four fragments (83, 28, 34 and 42 kDa). The C-terminal 42 kDa fragment (MSP-142) undergoes further processing to form MSP-133, which is shed, and MSP-119, which remains on the Mz surface and is taken into the newly invaded erythrocytes. Monoclonal antibodies to MSP-119 inhibit Mz invasion in vitro, and sera from P. falciparum immune adult humans and P. chabaudi immune mice revealed a major role for MSP-119 specific antibodies in mediating the invasion-inhibition (O'Donnell et al., 2001). The 19 kDa fragment is reported to be highly conserved in P. falciparum and contains a series of cysteine residues that are conserved among species of plasmodia infecting humans, primates and rodents (Daly et al., 1992). Different effector mechanisms of antibodies against MSP-1 are being suggested which primarily involve inhibition of erythrocyte invasion by Mzs (Tolle et al., 1993; Locher et al., 1996; O'Donnell et al., 2001; Vukovic et al., 2003), blocking the processing of larger mature MSP-1 protein on the Mz surface (Blackman et al., 1994) and through macrophage Fc receptors to induce ADCC (Bouharoun-Tayoun et al., 1995; Ravetch and Clynes, 1998). MSP-119-specific immunoglobulin IgG 3 monoclonal antibody can passively transfer protection to mice deficient in FcãRI receptors whose macrophages cannot bind IgG 3 (Vukovic et al., 2000). Studies in P. yoelii model by Wipasa et al. (2002) show that antibodies specific to MSP-119 alone can induce protective immunity and that effector T-cells specific to MSP-119 play no role in immunity. However, such antibodies must be produced during challenge. Specific cellular immune responses induced by MSP-1 can be protective against exoerythrocytic forms of P. yoelii (Kawabata et al., 2002). Banyal and Elangbam MSP-2 is encoded by a single gene and is a 4552 kDa integral membrane glycoprotein anchored on the surface of Mz by a glycosylphosphatidyl inositol (GPI) moeity (Weisman et al., 2001) and a target of host protective immune responses as monoclonal antibodies specific to MSP-2 have inhibited parasites growth in vitro. Mice immunized with conserved regions of P. falciparum MSP-2 have been protected against challenge with P. chabaudi. Antibodies to MSP-2, mainly IgG 3, have been detected in sera of people living in endemic areas. Human trials of multi subunit vaccines containing MSP-2 have been undertaken both in non-exposed and malaria-exposed individuals (Genton et al., 2000). MSP-3 is a secreted polymorphic antigen associated with erythrocytic schizonts and Mzs. P. falciparum MSP-3 has been shown to range from 40-76 kDa depending on the isolate and has been implicated in induction of ADCI (Audran et al., 2005; Druilhe et al., 2005). P. vivax MSP-3 is associated with but not anchored in the Mz membrane and is structurally related to P. falciparum MSP-3 and 140 kDa MSP of P. knowlesi (Galinski et al., 1999). P. falciparum MSP-4 is a 40 kDa protein containing a single epidermal growth factor (EGF)-like domain at the C-terminus that is synthesized at the late ring stage and transported to the parasite surface, anchored to the Mz membrane by a GPI moiety. Studies analyzed a region of chromosome 2 in P. falciparum and identified 3 clustered genes that encode GPI-anchored Mz surface proteins in tandem MSP-2, MSP-5 and MSP-4 and MSP-5 encoding a 40 kDa protein located on the Mz surface (Marshall et al., 1998). A homologue of P. falciparum MSP-4 and MSP-5 in P. chabaudi designated Pc MSP 4/5 encoding a protein of apparently 36 kDa contains a single EGF-like domain near the C-terminus. Murine homologue of MSP-4 induces protective immunity in mice against lethal challenge with P. yoelii. Anti-MSP-4 antibodies are highly prevalent and present at high level in individuals in malaria endemic area and are mainly IgG 1 and IgG 3. Proteolytic processing of MSP-1 precursor produces two components p 36 (MSP-636) and p 22 (MSP-722) which are associated with the shed MSP-1 complex. The 36 kDa protein is derived from a larger precursor MSP-6 and so designated as MPS-636. Antibodies against recombinant protein containing the C-terminal of MSP-6 36 bound to parasite surface or the parasitophorous vacuole within schizonts (Trucco et 21 Malaria vaccine development al., 2001). MSP-6 reactive antibodies are generated in a natural human infection and antibodies on an MSP-3 peptide cross reacted with MSP-6 which suggest it to be a target of ADCI. The 22 kDa protein designated MSP-722 is the result of protease cleavage of precursor MSP-7 expressed in mature schizonts (Pachebat et al., 2001). Another asexual stage parasite protein of P. falciparum containing two EGF-like domains near the C-termini is designated as MSP-8 (Black et al. 2001). Apical membrane antigen (AMA)-1: Apical membrane antigen is a trans-membrane protein present on the surface of Mzs and involved in the parasite invasion of erythrocytes. AMA-1 has been identified in all Plasmodia as a relatively conserved sequence and synthesized de novo as a 66 kDa protein except in P. falciparum and P. reichenowi. In P. falciparum and P. reichenowi, AMA-1 is a 83 kDa protein processed by proteolytic cleavage between different domains into a 66 kDa form (Pf AMA-166) which is further proteolytically cleaved and shed as soluble ectodomain (Howell et al., 2001). P. falciparum Pf 83/AMA-1 is the analogue of 66 kDa P. knowlesi protective Mz protein, Pk 66/AMA-1 that is expressed in late schizonts and localized in the Mz apex. The full length 83-kDa remains apically restricted while the processed 66-kDa becomes circumferentially associated with the Mz surface. An Escherichia coli expressed recombinant P. falciparum AMA-1 induced in vitro growth inhibitory anti-AMA1 antibodies which recognize both strain specific and conserved epitopes and show AMA-1 to be a natural target of protective antibody responses (Hodder et al., 2001). Recombinant AMA-1 proteins either alone or in combination with MSP-1 have also been evaluated for its efficacy against blood-stage malaria in animal models (Burns et al., 2003). Several of the potential malaria blood-stage vaccine candidate antigens are expressed on the rhoptries, apical organelles involved in erythrocyte invasion. Rhopty Asociated Protein (RAP-1) is synthesized as a 86 kDa precursor N-terminally cleaved to generate 82 kDa molecule, P 82, that is further processed to give a 68 kDa molecule, P 68 (Howard et al., 1998). Purified complexes of RAP-1 and RAP-2 used in experimental immunization of Saimiri monkeys have shown partial protection against P. falciparum infection, whereas anti-RAP monoclonal antibodies have shown inhibition of parasite replication in vitro (Ridley et al., 1990). Proteins involved in the targeting of particular erythrocyte subpopulation for Mz invasion have been intensely studied as a possible means of blocking Mz attachment to erythrocytes. In P. falciparum, glycophorin A is the erythrocyte receptor for Mzs whose major ligand is the erythrocyte-binding antigen-175 (EBA-175) located in the microneme. Blocking an EBA-175 binding site inhibits parasite multiplication in vitro and immunization of Aotus monkeys with recombinant EBA-175 region II induced anti-parasite effect (Jones et al., 2001). In P. vivax, Duffy antigen receptor for chemokines (DARC) is the receptor in erythrocyte for Mz which expresses Duffy-binding protein (DBP; Yazdani et al., 2005). P. vivax and P. knowlesi DBPs and their orthologue EBA175 bind well-defined glycoprotein motifs on erythrocyte membrane. P. vivax reticulocyte binding proteins (PvRBP-1 and PvRBP-2) attach to reticulocyte enriched erythrocytes. PvRBP-1 and PvRBP-2 have molecular masses of 325 kDa and 330 kDa, respectively, and share similar structures (Galinski et al., 2000). PvRBP-2 appears to be distantly related to a 235 kDa rhoptry protein of P. yoelii. P. falciparum genes homologous to P. vivax RBP-1 and -2 encoding high molecular mass proteins of > 300 kDa are expressed in late schizonts. Two PvRBP-2 orthologues of P. falciparum termed P. falciparum RBP-2 homologues a and b (Pf RBP 2-Ha and Pf RBP 2-Hb) along with RBP-2 of P. vivax and P. yoelii 235 kDa protein, constitute an important Plasmodium family important for Mz invasion. Parasitized erythrocytes Parasite antigens expressed on the infectederythrocytes are targets for antibodies to act on, and several such molecules have been identified. Ringinfected erythrocyte surface antigen (RESA) is one of the most studied with molecular weights 155 and 130 kDa, and anti-RESA antibodies raised in mice inhibit parasite growth in vitro (Chopra et al., 2000). RESA has also been used as a component in multiantigen blood-stage vaccine together with MSP-1 and-2 in Phase I clinical trials. The serine-rich protein (SERP/SERA) is a 120 kDa soluble protein expressed in the schizont stage, whereas glutamate rich protein (GLURP) is expressed during all stages of the parasite development in human host as a 220 kDa protein. Several immuno-epidemiological studies have identified high anti-Ro-GLURP IgG levels as predictors of protection against high parasitaemia and febrile malaria episodes (Soe et al., 2004). There is an association between protection against febrile malaria 22 and presence of anti Ro-GLURP antibodies in intense malaria transmission areas, and further that increasing levels of IgG 1 and IgG 3 antibodies are associated with reducing P. falciparum parasite densities (Lusingu et al., 2005). Erythrocyte membrane proteins (EMP) e. g. EMP 1, 2 and 3 are also located on erythrocyte membrane; however, Pf EMP-3 is expressed not only on erythrocyte surface but also by liver stage parasites and sporozoites (Gruner et al., 2001). Parasite toxins The toxic basis of malarial pathogenesis was first conjectured by Camillo Golgi in 1886 (Golgi, 1886). Malaria GPI is considered a candidate toxin as it induces cytokine like TNF-α and adhesion expression in macrophages and vascular endothelium which are associated with malaria pathogenesis. Antibodies to GPI lipid domains have been associated with protection against malaria. GPI is a highly conserved pro-inflammatory endotoxin of parasite origin and synthetic anti-GPI can be used as a plausible anti-toxic vaccine (Schofield et al. 2002). Sexual stage antigens Various studies have demonstrated that antibodies directed against the sexual-stage antigens can prevent fertilization in the mosquito thereby interrupting the transmission of malaria. Transmission blocking vaccines (TBVs) would reduce or interrupt malaria transmission in human and mosquito populations within a community as a whole but confer no protection to an individual recipient. Such a transmission blocking vaccine when given in combination with a pre-erythrocytic or blood-stage vaccine would prevent or reduce the spread of parasites which become resistant to such vaccines and would thus prolong the effective life of other malaria vaccines. Antigens expressed on the surface of sexual stages i.e. gametocyte, gamete, zygote and ookinete of malaria parasite are being considered as promising targets for developing a transmission blocking vaccine. These vaccines induce antibodies in human host that inhibit parasite development within mosquito vector, thus, blocking parasite transmission. Cloning of genes and subsequent recombinant proteins have shown to induce transmission-blocking antibodies in animal models. Two groups of antigens have been explored to block the propagation of sexual parasites. (i) Pfs 48/45 and Pfs 230 antigens. These proteins belong to a family unique to Plasmodium with Banyal and Elangbam characteristic arrangement of cysteine containing domains within the proteins. Monoclonal antibodies against these proteins have been effective in blocking the infectivity of the parasites to mosquitoes (Templeton and Kaslow, 1999). (ii) Ps 25 and Ps 28 antigens. These are surface proteins expressed on the zygotes and mature ookinetes of malaria parasite. The ookinete surface proteins of P. falciparum, Pfs 25 and Pfs 28, are target antigens for a possible vaccine and their homologous proteins have been cloned from P. vivax and other species. P. falciparum Pfs 25 has been tested in Phase I clinical trials in human volunteers (Kaslow and Shiloach, 1994). Yeast produced recombinant Pvs 25 and Pvs 28 are highly immunogenic and antisera recognized corresponding molecules expressed by field-isolated parasites in Thailand (Sattabongkot et al., 2003). CURRENT STATUS AND PROSPECTS OF MALARIA VACCINES The development of a malaria vaccine remains an urgent need for most people living in malaria endemic regions. Two lines of evidence suggest the feasibility of a malaria vaccine: firstly, immunity can be acquired as a result of natural exposure to infection, and secondly, numerous experimental malaria infections in animal models and human volunteers can be protected through various immunization strategies. The distinct developmental stages of malaria parasite provide numerous targets for vaccine development. Because the production of live, attenuated or killedinactivated malaria vaccine is not practical, the aim has been to develop sub-unit vaccines. In this, part or complete antigens are identified from the pathogen's proteomic complement, which can induce protective immunity to the whole pathogen on vaccination. The general target for a subunit vaccine has been to identify critical target antigens at each stage of the lifecycle of malaria parasite. Another strategy is to assemble peptide sequences from a range of parasite antigens into different combinations that are then tested for immunogenicity in animal models and human volunteers. In the new generation of DNAbased subunit vaccines, DNA sequences from P. falciparum parasites have been inserted into plasmid DNA molecules (DNA vaccines) or various recombinant attenuated DNA viruses (recombinant viral vaccines) to generate candidate vaccines (Wang et al., 1998). DNA vaccines are taken up by host cells, protein expressed and T-cells are primed to form memory T-cell populations while in recombinant viral Malaria vaccine development vaccines, cells are actively infected and express the recombinant malaria proteins. Both vaccines induce high levels of effector T-cell immune responses which in combination with antibody responses are both critical in protection against malaria. Three stages in the life-cycle of malaria parasite are targeted for vaccine development (Table I). Pre-erythrocytic vaccines: These would prevent the disease by targeting sporozoites or schizont infected liver cells and by preventing the release of primary Mzs from infected hepatocytes. Such a vaccine that successfully interferes with pre-erythrocytic development would terminate infections before they had any chance of causing clinical illness and would be beneficial to travellers and residents of malaria endemic countries. Numerous vaccine constructs have been developed with CS protein being the prime target. RTS, S/ASO 2A: This vaccine candidate is the most advanced in clinical development and is based on the CS protein of P. falciparum comprising of two polypeptides RTS and S expressed in yeast. RTS is a single polypeptide chain comprising of CS protein fused to hepatitis B surface antigen (HBsAg), while S is a polypeptide of HBsAg. The two polypeptides assemble to form composite particulate structure (RTS, S) that constitute the vaccine antigen. This vaccine when administrated with the adjuvant ASO2A, an oil in water emulsion consisting of the immunostimulants monophosphoryl lipid A and saponin derivative QS 21, has shown good immunogenicity and efficacy in clinical trials. Phase II safety and immunogenicity trials in malaria naïve and malaria-immune subjects have shown it to be safe and immunogenic and conferred 50% sterile immunity in volunteers but immunity waned with time lasting only up to six months in a few cases. Phase I field evaluation in malaria experienced adults was done in Gambia (Doherty et al., 1999). Later a randomized, double-blind controlled Phase IIb efficacy study was carried out in malaria-experienced adult Gambian men that gave overall efficacy of 34% (Bojang et al., 2001). Efficacy trial in children in Mozambique and Phase I trials in children in The Gambia gave encouraging results. A Phase IIb proof of concept efficacy study was done in children in Southern Mozambique which showed that the vaccine was highly immunogenic and had an efficacy of 58% against severe malaria (Alonso et al., 2004). ICC/-1132 CS/hepatitis B core particle: This is a 23 CS-based particle vaccine that uses the highly immunogenic hepatitis B core antigen (HBcAg) as a delivery platform. It includes T-cell epitopes and Bcell epitopes and is engineered so that the B-cell epitopes are exposed on the surface of virus-like particles (VLPs), whereas the T-cell epitopes are within the interior. Three Phase I clinical trials in healthy, malaria-naïve adults have been undertaken. Phase I trials in UK and Germany evaluated aluminum hydroxide and Montanide ISA 720 formulations safe and well tolerated (Walther et al., 2005). Multi-epitope TRAP: This vaccine construct consists of plasmid DNA or recombinant attenuated live viral vector like adenovirus, fowl pox and modified vaccinia Ankara (MVA) in a prime-boost strategy, the most advanced of which is a multiepitope (ME) string fused to TRAP. The ME portion contains two B-cell, 14 CD8+ and three CD4+ epitopes from six sporozoite and/or liver stage antigens, including CS, LSA-1 and LSA-3. Several phase I and Phase IIb sporozoite challenge studies have been conducted in malaria-naïve adults in UK which is reported to induce high T-cell responses (McConkey et al., 2003). Phase I safety trials were investigated in adults and children in The Gambia also (Moorthy et al., 2003) and a further Phase IIb proof of concept trial is being conducted in The Gambia. Plasmid DNA vaccines: Intense efforts have been made to develop effective DNA-based vaccines to the liver and blood stages with combinations of plasmid DNA vaccines targeting one or more P. falciparum antigens. Several single and combination vaccines, including CS alone and CS with SSP2/TRAP, LSA-1, EXP-1 and LSA-3 (MuStDO-5) have undergone Phase I and IIa clinical trials (Doolan and Hoffman, 2001). DNA vaccines require viral boosting to induce strong T-cell immunogenicity. Asexual blood-stage vaccines: A second strategy for vaccine development is to target immune responses against the asexual blood stages with the aim to prevent or contain the disease process by suppressing parasite replication either by preventing Mz invasion of erythrocytes or by attacking parasite inside host erythrocytes. A blood stage vaccine would be effective in reducing mortality in endemic countries. The principal target of asexual stage vaccine is the Mz with the major target proteins like MSP-1, 2, 3 and AMA-1. A vaccine based on the C-terminal of P. falciparum MSP-1 consisting of a 42 kDa protein produced as a lyophilized recombinant antigen expressed in E. coli 24 Banyal and Elangbam Table I: Malaria vaccines in clinical trials Stages Vaccine/antigen Research group Trial phase Pre-erythrocytic Stage RTS,S: Hybrid P. falciparum CSPHBsAg particles + AS02 adjuvant CSP C-ter peptide + Montanide ISA 720 ICC-1132: Hybrid CSP multiepitope-HBc VLPs DNA vaccines (including MuStDO-5: CSP/LSA-1/ LSA-3/EXP1/TRAP) Live recombinant FPV- or MVA-CSP + LSA-1 epitope Live recombinant MVAmultiepitope string + TRAP Live recombinant Ad-CSP Other live recombinant vaccines (MVA, cold-adapted influenza virus)-CSP LSA-3 (long peptides; lipopeptide;recombinant) LSA-1, SALSA, other liver-stage antigens GSK / WRAIR/MVI Phase IIb Dictagen / Lausanne Univ Phase Ib Apovia/MVI Phase II US Navy/ Vical Phase I Oxford Univ/ Oxxon/MVI Phase Ib Oxford Univ/Oxxon Phase Ib Crucell/GSK/WRAIR/NIAID Oxford Univ; NYU Phase Ia Preclinical Pasteur Institute/WRAIR/ GSK Phase Ia Hawaii Biotech; Epimmune Preclinical GSK/WRAIR/MVI MVDU; NIAID Sec Military Med Univ Shanghai/ Wanxing harmaceuticals/WHO Phase Ib/ II Phase Ib Phase I NIAID; Hawaii Biotech; AECOM; Univ Maryland Preclinical to Phase I Pasteur Institute/ AMANET/EMVI EMVI/SSI EMVI/SSI Phase Ib Phase I Phase I Monash Queensland Med Res Institute/ WEHRI Osaka Univ/Biken Various groups Preclinical Phase II Phase I Preclinical NIH NIH Phase I Preclinical Asexual erythrocytic stage Sexual stage MSP-1 42 kD + AS02 AMA-1 PfCP 2.9: MSP-1-AMA-1 fusion protein (yeast) + Montanide ISA 720 Other MSP-1 derivatives (peptides, Salmonella or BCG recombinants) MSP-3 long peptides GLURP long peptide MSP-3-GLURP hybrid long peptide + Montanide ISA 720 MSP-4, -5 Combination B: MSP-1, -2, RESA + Montanide SE36 Other blood-stage antigens (EBA-175,EBP2, MAEBL, RAP-2, EMP-1, DBL-á..) PfS25 (yeast) PvS25 and other sexual-stage antigens Source: www.who.int/vaccine_research/documents/en/status_table.pdf. 25 Malaria vaccine development has shown great promise (Angov et al., 2003). This vaccine reconstituted with ASO2A, falciparum Mz protein 1 (FMP-1)/ASO2A was found to be safe and immunogenic in mice, rabbits and rhesus macaques. The safety and immunogenicity trial has been conducted in malaria-naïve individuals. A Phase I safety study in malaria-experienced Kenyan adults has been completed and is scheduled to begin a Phase I study in children in malaria-endemic region of western Kenya. A chimeric fusion of domain III of AMA-1 and the 19kDa portion of MSP-1 called P. falciparum chimeric protein 2 (Pf CP-2.9) containing conserved portions of both proteins is also being developed (Genton and Corradin, 2002). Pf CP-2.9 expressed in Pichia pastoris has demonstrated good immunogenicity to both portions of antigen in mice, rabbits and nonhuman primates. A preclinical trial showed its effectiveness, and a Phase I trial in malaria-naïve healthy adults with Montanide ISA720 formulation is under way that began in February 2006. The idea of a multi-component vaccine against bloodstage parasites was approached by Patarroyo and colleagues with a synthetic peptide vaccine SPf66, containing sequence from MSP-1 and two other blood-stage antigens combined with CS protein (Patarroyo et al., 1987). The vaccine underwent four Phase III clinical trials in different locations. Studies in Colombian adults and children (Valero et al., 1993) and in Tanzanian children (Alonso et al., 1994) gave encouraging results but no significant protection was observed in studies in Gambian infants (D'Alessandro et al., 1995) and in Thailand (Nosten et al., 1996). Sexual-stage vaccines: The third vaccine strategy is the development of a sexual-stage vaccine. Antigens of gametocyte 48/45 kDa and 230 KDa and the 25 kDa and 28 kDa post-fertilization antigens are leading candidates for transmission blocking vaccine (TBV). Pvs 25 adjuvanted with aluminum hydroxide is currently in Phase I clinical trial. Pvs 25 adjuvanted with cholera toxin (CT) when used to immunize intranasally induced significant Th-2 type immune response in mice and the antisera completely blocked parasite transmission (Arakawa et al., 2003). CONCLUSIONS Immunity to malaria is complex and not clearly defined with the parasite exhibiting various immunoevasive mechanisms like antigenic variation, polymorphism and polyclonal B-cell activation. Protection in endemic areas takes years to develop and there is no clearly defined immune response that can effectively protect against the disease. There is also the need to develop novel adjuvants that can enhance the cell-mediated immunity, and also for the development of novel vectors and vaccine formulations that will produce optimal protective immune responses. Some of the recent vaccine trials like RTS, S which showed 58% efficacy against severe disease have given encouraging results. Malaria vaccine development is at an exciting stage with an optimistic outlook. Recent advances in understanding the immune mechanisms and the volume of information generated from malaria parasite genome would help in better understanding and identification of critical antigens for vaccine development. Perhaps, given the complexity of the parasite, vaccine based on all or many antigens would be more effective than the one based on a single antigen. ACKNOWLEDGMENTS Ms. N. Elangbam is grateful to University Grants Commission, New Delhi, for financial assistance in the form of a Junior Research Fellowship (National Eligibility Test). REFERENCES Alonso PL, Sacarlal J, Aponte JJ, Leach A, Macete E, Milman J, Mandomando I, Spiessens B, Guinovart C, Espasa M, Bassat Q, Aide P, Ofori-Anyinam O, Navia MM, Corachan S, Ceuppens M, Dubois MC, Demoite MA, Dubovsky F, Menendez C, Tornieporth N, Ballou WR, Thompson R and Cohen J. 2004. Efficacy of the RTS, S/AS02A vaccine against Plasmodium falciparum infection and disease in young African children: randomized controlled trial. Lancet 364: 1411-1420. Alonso PL, Smith T, Schellanberg JR, Masanja H, Mwankuyse S, Urassa H, de Azevedo IB, Chongela J, Kobero S, Menendez C, Hurt N, Thomas MC, Lyimo E, Weiss NA, Hayes R. Kitva AY, Lopez MC, Kilama WL, Teuscher T and Tanner M. 1994. Randomised trial of efficacy of SPf66 vaccine against Plasmodium falciparum malaria in children in Southern Tanzania. Lancet 344: 1175-1181. Angov E, Aufiero BM, Turgeon AM, Van Handenhove M, Ockenhouse CF, Kester KE, Walsh DS, McBride JS, Dubois MC, Cohen J, Haynes JD, Eckels KH, Heppner DG, Ballou WR, Diggs CL and Lyon JA. 2003. Development and preclinical analysis of a Plasmodium falciparum Mz surface protein-1 (42) malaria vaccine. Mol Biochem Parasitol 128: 195-204. Arakawa T, Tsuboi T, Kishimoto A, Sahabongkot J, Suwanabun N, Rungruang T, Matsumoto Y, Tsuji N, Hisaeda H, Stowers A, Shimabukuro I, Sato Y and Torii M. 2003. Serum 26 antibodies induced by intranasal immunization of mice with Plasmodium vivax Pvs 25 co-administered with cholera toxin completely block parasite transmission to mosquitoes. Vaccine 21: 3143-3148. Artavanis-Tsakonas K, Tongren JE and Riley EM. 2003. The war between the malaria parasite and the immune system : immunity, immunoregulation and immunopathology. Clin Exp Immunol 133: 145-152. Audran R, Cachat M, Lurati F, Soe S, Leroy O, Corradin G, Druilhe P and Spertini F. 2005. Phase 1 malaria vaccine trial with a long synthetic peptide derived from the Mz surface protein 3 antigen. Infect Immun 73: 8017-8026. Banyal HS and Inselburg J. 1985. Isolation and characterization of parasite inhibitory Plasmodium falciparum monoclonal antibodies. Am J Trop Med Hyg 34: 1055-1064. Bastianelli G and Bignami A. 1899. Sullo sviluppo dei parassiti della terzana nell' Anopheles claviger. Atti Soc Studi Malar 1: 28-49. Black CG, Tieqiao W, Wang L, Hibbs AR and Coppel RL. 2001. Mz surface protein 8 of Plasmodium falciparum contains two epidermal growth factor-like domains. Mol Biochem Parasitol 114: 129-262. Blackman MJ, Scott-Finningan TJ, Shai S and Holder AA. 1994. Antibodies inhibit the protease-mediated processing of a malaria Mz surface protein 1. J Exp Med 180: 389-393. Bojang KA, Milligan PJM, Pinder M, Vigneron L, Alloueche A, Kester KE, Ballou WR, Conway DJ, Reece WHH, Gothard P, Yamuah L, Delchambre M, Voss G, Greenwood BM, Hill AVS, McAdam KPWJ, Tornieporth N, Cohen J and Doherty T. 2001. Efficacy of RTS, S/AS02A malaria vaccine against Plasmodium falciparum infection in semi-immune adult men in The Gambia: a randomized trial. Lancet 358: 19271934. Bouharoun-Tayoun H, Oeuvray C, Lunel F and Druilhe P. 1995. Mechanisms underlying the monocyte-mediated antibodydependent killing of Plasmodium falciparum asexual blood stages. J Exp Med 182: 409-418. Boyd MF. 1949. 'Malariology'. Saunders, Philadelphia, Pennsylvania. Burns JM Jr, Flaherty PR, Romero MM and Weidanz WP. 2003. Immunization against Plasmodium chabaudi malaria using combined formulations of apical membrane antigen-1 and Mz surface protein-1. Vaccine 21: 1843-1852. Cavacini L, Parke LA and Weidanz WP. 1990. Resolution of acute malarial infections by T-cell dependent non-antibody mediated mechanisms of immunity. Infect Immun 58: 29462950. Chelimo K, Ofulla A, Narum DL, Kajura JW, Lanar DE and John CC. 2005. Antibodies to Plasmodium falciparum antigens vary by age and antigenin children in a malariaholoendemic area of Kenya. Pediatric Infect Dis J 24: 680684. Banyal and Elangbam Chopra N, Biswas S, Thomas B, Sabhnani L and Rao DN. 2000. Inducing protective antibodies against ring-infected erythrocyte surface peptide antigen of Plasmodium falciparum using immunostimulating complex (ISCOMS) delivery. Med Microbiol Immunol 189: 75-83. Couper KN, Phillips RS, Brombacher F and Alexander J. 2005. Parasite-specific IgM plays a significant role in the protective immune response to asexual erythrocytic stage Plasmodium chabaudi AS infection. Parasit Immunol 27: 171-180. D'Alessandro U, Leach A, Drakeley CJ, Bennett S, Olaleye BD, Pegan GW, Jawara M, Langerock P, George MO, Targett GA and Greenwood BM. 1995. Efficacy trial of malaria vaccine SPf 66 in Gambian infants. Lancet 346: 462-467. Daly TM, Burns Jr, JM and Long CA. 1992. Comaprison of the carboxy-terminal, cysteine-rich domain of the Mz surface protein-1 from several strains of Plasmodium yoelii. Mol Biochem Parasitol 52: 279-282. Daubersies P, Thomas AW, Millet P, Brahimi K, Langermans JAM, Ollomo B, Benmohamed L, Slierendreg, B, Eling W, Van Belkum A, Dubreuil G, Meis JFGM, Guerin-Marchand C, Cayphas S, Cohen J, Gras-Masse H and Druilhe P. 2000. Protection against Plasmodium falciparum malaria in chimpanzees by immunization with the conserved preerythrocytic liver stage antigen 3. Nature Med 6: 12581263. Dodoo DF, Omer F, Todd J, Akanmor B, Koram K and Riley EM. 2002. Absolute levels and ratios of pro-inflammatory and anti-inflammatory cytokine production in vitro predict clinical immunity to P. falciparum malaria. J Infect Dis 185: 971-979. Doherty JF, Pinder M, Tornieporth N, Carton C, Vigneron L, Milligon P, Ballou WR, Holland CA, Kester KE, Voss G, Momin P, Greenwood BM, McAdam KP, and Cohen J. 1999. A phase I safety and immunogenicity trial with the candidate malaria vaccine RTS, S/SBAS 2 in semi-immune adults in The Gambia. Am J Trop Med Hyg 61: 865-868. Doolan DL and Hoffman SL. 1999. NK Cells and IL-12 are required for antigen-specific adaptive immunity against malaria initiated by CD8+ T cells. J Immunol 163: 884-892. Doolan DL and Hoffman SL. 2000. The complexity of protective immunity against liver-stage malaria. J Immunol 165: 1453-1462. Doolan DL and Hoffman SL. 2001. DNA-based vaccines against malaria: status and promise of the Multi-Stage Malaria DNA vaccine Operation. Int J Parasitol 31: 753762. Druilhe P, Spertini F, Soesoe D, Corradin G, Mejia P, Singh S, Audran R, Bouzidi A, Oeuvray C and Roussilhon C. 2005. A malaria vaccine that elicits in humans antibodies able to kill Plasmodium falciparum. PLOS Med 2: e344. Egan JE, Weber JL, Ballou WR, Hollingdale MR, Majarian WR, Gordon DM, Maloy WL, Hoffman SL, Wirtz RA, Schneider I, Woollett GR, Young JF and Hockmeyer WT. Malaria vaccine development 27 1987. Efficacy of murine malaria sporozoite vaccines: implications for human vaccine development. Science 236: 453-456. Herrera M. 2004. Use of long synthetic peptides to study the antigenicity and immunogenicity of the Plasmodium vivax circumsporozoite protein. Int J Parasitol 34: 1535-1546. Galinski MR, Corredor-Medina C, Povoa M, Crosby J, Ingravallo P and Barnwell JW. 1999. Plasmodium vivax Mz surface protein-3 contains coiled-coilmotifs in an alaninerich central domain. Mol Biochem Parasitol 101: 131-147. Hodder AN, Crewther PE and Anders RF. 2001. Specificity of the protective antibody response to apical membrane antigen 1. Infect Immun 69: 3286-3294. Galinski MR, Mengyao X and Barnwell JW. 2000. Plasmodium vivax reticulocyte binding protein-2 (Pv RBP2) shares structural features with Pv RBP-1 and the Plasmodium yoelii 235 kDa rhoptry protein family. Mol. Biochem Parasitol 108: 257-262. Genton B and Corradin G. 2002. Malaria vaccines: from the laboratory to the field Curr. Drug Targets Immune Endocrinol Metabol Disord 2: 255-267. Genton B, Al-Yaman F, Anders R, Saul A, Brown G, Pye D, Irving DO, Briggs WRS, Mai A, Ginny M, Adiguma T, Rare L, Giddy A, Reberliske R, Stuerchler D and Alpers MP. 2000. Safety and immunogenicity of a three-component blood-stage malaria vaccine in adults living in an endemic area of Papua New Guinea. Vaccine 18: 2504-2511. Golgi C. 1886. Sull infezione malarica. Arch Sci Med.,(Torino) 10: 109-135. Good MF and Doolan DL. 1999. Immune effector mechanisms in malaria. Curr Opin Immunol 11:412-419. Grassi B and Feletti R. 1892. Contribuzione allo studio dei parassiti malarici. Atti Accad Gioenia Sci Nat Catania, 16: 16-20. Grassi B, Bignami A and Bastianelli G. 1899a. Ciclo evolutivo delle semilune nell' Anopheles claviger. Atti. Soc. Studi. Malar 1: 14. Grassi B, Bignami A and Bastianelli G. 1899b. Resoconto degli studi fatti sulla malaria durante il mese di Gennaio. Atti. Accad Naz Lencei Cl Sci Pis Nat Nat Rend 8: 100-104. Grun JL and Weidanz WP. 1981. Immunity to Plasmodium chabaudi adami in the B-cell deficient mouse. Nature 290: 143-145. Gruner AC, Brahimi K, Eling W, Konings R, Meis J, Aikawa M, Daubersies P, Guerin-Marchand C, Mellouk S, Snounou G and Druilhe P. 2001. The Plasmodium falciparum knobassociated Pf EMP 3 antigen is also expressed at preerythrocytic stages and induces antibodies which inhibit sporozoite invasion. Mol Biochem Parasitol 112: 253-261. Herrera S, Bonelo A, Perlaza BL, Farnandez OL, Victoria L, Lenis AM, Soto L, Hurtado H, Acuna LM, Velez JD, Palacios R, Chen-Mok M, Corradin G and Arevalo-Herrera M. 2005. Safety and elicitation of humoral and cellular responses in Colombian malaria-naive volunteers by a Plasmodium vivax circumsporozite protein derived synthetic vaccine. Am J Trop Med Hyg 73: 3-9. Herrera S, Bonelo A, Perlaza BL, Valencia AZ, Cifuentes C, Hurtado S, Quintero G, Lopez JA, Corradin G and Arevalo- Howard RF, Jacobson KC, Rickel E and Thurman J. 1998. Analysis of inhibitory epitopes in the Plasmodium falciparum rhoptry protein RAP-1 including identification of a second inhibitory epitope. Infect Immun 66: 380-386. Howell SA, Withers-Martinez C, Kocken CH, Thomas AW and Blackman MJ. 2001. Proteolytic processing and primary structure of Plasmodium falciparum apical membrane antigen-1 (Pf AMA-1). J Biol Chem 276: 31311-31320. Jones TR, Narum DL, Gozalo AS, Aguiar J, Fuhrmann SR, Liang H, Haynes JD, Moch JK, Lucas C, Luu T, Magill AJ, Hoffman SL and Sim BKL. 2001. Protection of Aotus monkeys by Plasmodium falciparum EBA-175 region II DNA prime-protein boost immunization regimen. J Infect Dis 183: 303-312. Kaslow DC and Shiloach J. 1994. Production, purification and immunogenicity of a malaria transmission blocking vaccine candidate: TBV 25H expressed in yeast and purified using nickel-NTA agarose. Biotechnology 12: 494-499. Kawabata Y, Udono H, Honma K, Ueda M, Mukae H, Kadota J, Kohno S and Yui K. 2002. Mz surface protein-1 specific immune response is protective against exoerythrocytic farms of Plasmodium yoelii. Infect Immun 70: 6075-6082. Kremsner PG, Winkler X, Wilding E, Prada J, Bienzle U, Graninger W and Nussler A. 1996. High plasma levels of nitrogen oxides are associated with severe disease and correlate with rapid parasitological and clinical cure in Plasmodium falciparum malaria. Trans R Soc Trop Med Hgy 90: 44-47. Locher CP, Tam LQ, Chang SP, McBride JS and Siddiqui WA. 1996. Plasmodium falciparum: gp 195 tripeptide repeatspecific monoclonal antibody inhibits parasite growth in vitro. Exp Parasitol 84: 74-83. Lusingu JPA, Vestergaard LS, Alifrangis M, Mmbando BB, Theisen M, Kitua A, Lemnge MM and Theander TG. 2005. Cytophilic antibodies to Plasmodium falciparum glutamate rich protein are associated with malaria protection in an area of holoendemic transmission. Malaria J 4: 48-55. MacMicking J, Xie QW and Nathan C. 1997. Nitric oxide and macrophage function. Annu Rev Immunol 15: 323-350. Marshall VM, Tieqiao W and Coppel RL. 1998. Close linkage of three Mz surface protein genes on chromosome 2 of Plasmodium falciparum. Mol Biochem Parasitol 94: 13-25. McConkey SJ, Reece WH, Moorthy VS, Webster D, Dunachie S, Butcher G, Vuola JM, Blanchard TJ, Gothard P, Watkins K, Hannan CM, Everaere S, Brown K, Kester KE, Cummings J, Williams J, Heppner DG, Pathan A, Flanagan K, Arulanantham N, Roberts MT, Roy M, Smith GL, 28 Schneider J, Peto T, Sinden RE, Gilbert SC and Hill AV. 2003. Enhanced T-cell immunogenicity of plasmid DNA vaccines boosted by recombinant modified vaccinia virus Ankara in humans. Nature Med 9: 729-735. Meraldi V, Romero JF, Kensil C and Corradin G. 2005. A strong CD8+ T cell response is elicited using the synthetic polypeptide from the C-terminus of the circumsporozoite protein of Plasmodium berghei together with the adjuvant QS-21: quantitative and phenotypic comparison with the vaccine model of irradiated sporozites. Vaccine 23: 28012812. Moorthy VS, Pinder M, Reece WH, Watkins K, Atabani S, Hannan C, Bojang K, McAdam KP, Schneider J, Gilbert S, and Hill AV. 2003. Safety and immunogenicity of DNA/modified vaccinia virus Ankara malaria vaccination in African adults. J Infect Dis 188: 1239-1244. Muller HM, Reckmann I, Hollingdale MR, Bujard H, Robson KJ and Crisanti A. 1993. Thrombospondin related anonymous protein (TRAP) of Plasmodium falciparum binds specifically to sulfated glycocongugates and to HepG2 hepatoma cells suggesting a role for this molecule in sporozoite invasion of hepatocytes. EMBO J 12: 28812889. Nosten F, Luxemburger C, Kyle DE, Ballou WR, Wittes J, Wah E, Chongsuphajaisiddhi T, Gordon DM, White NJ and Sadoff JC. 1996. Randomised double-blind placebo controlled trial of SPf 66 malaria vaccine in children in North West Thailand. Lancet 348: 701-707. Nussenzweig RS, Vanderberg J. Most H and Orton C. 1967. Protective immunity produced by the injection of Xirradiated sporozoites of Plasmodium berghei. Nature 216: 160-162. Banyal and Elangbam Immunol., Karger, Basel.80, pp. 229-242. Plebanski M and Hill AV. 2000. The immunology of malaria infection. Curr Opin Immunol. 12: 437-441. Polley SD, Tetteh KKA, Cavanagh DR, Pearce RJ, Lloyd JM, Bojang KA, Okenu MN, Greenwood BM, McBride JS and Conway DJ. 2003. Repeat sequences in block 2 of Plasmodium falciparum Mz surface proteins are targets of antibodies associated with protection from malaria. Infect Immun 71: 1833-1842. Pombo DJ, Lawrence G, Hirunpetcharat C, Rzepczyk C, Bryden M, Clooman N, Anderson K, Mahakunkijcharoen Y, Martin LB, Wilson D, Elliott S, Eisen DP, Weinberg JB, Saul A and Good MF. 2002. Immunity to malaria after administration of ultra-low doses of red cells infected with Plasmodium falciparum. Lancet 360: 610-617. Potocnjak P, Yoshida N, Nussenzweig RS and Nussenzweig V. 1980. Monovalent fragements (Fab) of monoclonal antibodies to a sporozoite surface antigen (Pb 44) protect mice against malaria infection. J Exp Med 151: 1504-1513. Pouvelle B, Spiegel R, Hsiao L, Howard RJ, Morris RL, Thomas AP and Taraschi TF. 1991. Direct access to serum macromolecules by intraerythrocytic malaria parasites. Nature 353: 73-75. Ravetch JV and Clynes RA. 1998. Divergent roles for Fc receptors and complement in vivo. Annu Rev Immunol 16: 421-432. Ridley RG, Takacs B, Etlinger H and Scaife JG. 1990. A rhoptry antigen of Plasmodium falciparum is protective in Saimiri monkeys. Parsitology 101: 187-192. O'Donnell RA, de Koning-Ward TF, Burt RA, Bockarie M, Reeder C, Cowman AF and Crabb BS. 2001. Antibodies against Mz surface protein (MSP)-119, are a major component of the invasion-inhibitory response in individuals immune to malaria. J Exp Med 193: 1403-1412. Sattabongkot J, Tsuboi T, Hisaeda H, Tachibana M, Suwanabun N, Rongruang T, Cao Y, Stowers AW, Sirichaisinthop J, Coleman RE and Torii M. 2003. Blocking of transmission to mosquitoes by antibody to Plasmodium vivax malaria vaccine candidates Pvs 25 and Pvs 28 despite antigenic polymorphism in field isolates. Am J Trop Med Hyg 69: 536-541. Omer FM, Kurtzhals JAL and Riley EM 2000. Maintaining the immunological balance in parasite infections: a role for TGF-α ? Parasitol Today 16: 18-23. Schofield L, Hewitt MC, Evans K, Slomos MA and Seeberger PH. 2002. Synthetic GPI as a candidate antitoxic vaccine in a model of malria. Nature 418: 785-789. Pachebat JA, Ling IT, Grainger M, Trucco C, Howell S, Fernandez-Reyes D, Gunaratne R. and Holder AA. 2001. The 22 kDA component of the protein complex on the surface of Plasmodium falciparum Mzs is derived from a larger precursor, Mz surface protein 7. Mol Biochem Parasitol 117: 83-89. Seixas E, Fonseca L and Langhorne J. 2002. The influence of gamma delta T cells on the CD4+ T cells and antibody response during a primary Plasmodium chabaudi chabaudi infection in mice. Parasite Immunol 24: 131-140. Patarroyo ME, Romero P, Torres ML, Clavijo P, Moreno A, Martinzez A, Rodriguez R, Guzman F and Cabezas E. 1987. Induction of protective immunity against experimental infection with malaria using synthetic peptides. Nature 328: 629-632. Perlmann P and Troye-Blomberg M. 2002. Malaria and the immune system in humans. In: Malaria Immunology. Perlmann P and Troye-Blomberg M. (Edts.) Chem. Soe S, Theisen M, Roussilhon C, Aye KS and Druilhe P. 2004. Association between protection against clinical malaria and antibodies to Mz surface antigens in an area of hyperendemicity in Myanmar: complementarity between responses to Mz surface protein 3 and the 220 kilodalton glutamate-rich protein. Infect Immun 72: 247-252. Stephens JWW. 1922. A new malaria parasite of man. Ann. Trop Med Parasitol 16: 383-388. Tebo AE, Kremsner PG and Luty AJ. 2001. Plasmodium Malaria vaccine development falciparum: a major role for IgG3 in antibody-dependent monocyte-mediated cellular inhibition of parasite growth in vitro. Exp Parasitol 98: 20-28. Templeton TJ and Kaslow DC. 1999. Identification of additional members define a Plasmodium falciparum gene superfamily which includes Pfs 48/45 and Pfs 230. Mol Biochem Parasitol 101: 223-227. Tolle R, Fruh K, Doumbo O, Koita O, N'Diaye M, Fischer A, Dietz K and Bujard H. 1993. A prospective study of the association between the human humoral immune response to Plasmodium falciparum blood stage antigen gp 190 and control of malaria infections. Infect Immun 61: 40-47. Trucco C, Fernandez-Reyes D, Howell S, Stafford WH, ScottFinnigan TJ, Grainger M, Ogun SA, Taylor WR and Holder AA. 2001. The Mz surface protein 6 gene codes for a 36 kDa protein associated with the Plasmodium falciparum Mz surface protein-1 complex. Mol Biochem Parasitol 112: 91101. Tsuji M and Zavala F. 2003. T cells as mediators of protective immunity against liver stages of Plasmodium. Trends Parasitol 19: 88-93. Valero MV, Amador LR, Galindo C, Figuerao J, Bello MS, Murillo LA, Mora AL, Patarroyo G, Rocha CL, Rojas M, Aponte JJ, Sarmiento LE, Losada DM, Coronell CG, Ortega NM, Rosas JE, Alonso PL and Patarroyo ME. 1993. Vaccination with SPf66, a chemically synthesised vaccine. against Plasmodium falciparum malaria in Colombo. Lancet 341: 705-710. Vukovic P, Chen K, Qin Liu X, Foley M, Boyd A, Kaslow D and Good MF. 2003. Single-chain antibodies produced by phage display against the C-terminal 19 kDa region of Mz surface protein-1 of Plasmodium yoelii reduce parasite growth following challenge. Vaccine 20: 2826-2835. Vukovic P, Hogarth PM, Barnes N, Kaslow DC and Good MF. 2000. Immunoglobulin G3 antibodies specific for the 19kilodalton carboxyl-terminal fragment of Plasmodium yoelii Mz surface protein 1 transfer protection to mice deficient in Fc-ãRI receptors. Infect Immun 68: 3019-3022. Walther M, Dunachie S, Keating S, Vuola JM, Berthoud T, Schmidt A, Maier C, Andrews L, Andersen RF, Gilbert S, Poulton I, Webster D, Dubovsky F, Tierney E, Sarpotdar P, Correa S, Huntcooke A, Butcher G, Williams J, Sinden RE, 29 Thornton GB and Hill AVS. 2005. Safety, immunogenicity and efficacy of a pre-erythrocytic malaria candidate vaccine, ICC-1132 formulated in Seppic ISA 720. Vaccine 23: 857-864. Wang R, Charoenvit Y, Corradin G, Porozzi R, Hunter RL, Glenn G, Alving CR, Church P and Hoffman SL. 1995. Induction of protective polyclonal antibodies by immunization with a Plasmodium yoelii circumsporozoite protein multiple antigen peptide vaccine. J Immunol 154: 2784-2793. Wang R, Doolan DL, Le TP, Hedstrom RC, Coonan KM, Charoenvit Y, Jones TR, Hobart P, Margalith M, Ng J, Weiss WR, Sedegah M, de Taisne C, Norman JA and Hoffman SL. 1998. Induction of antigen-specific cytotoxic T lymphocytes in humans by a malaria DNA vaccine. Science 282: 476-480. Webster DP, Dunachie S, Vuola JM, Berthoud T, Keating S, Laidlaw SM, McConkey SJ, Poulton I, Andrews L, Andersen RF, Byon P, Butcher G, Sinden R, Skinner MA, Gilbert SC and Hill VS. 2005. Enhanced T cell mediated protection against malaria in human challenges by using the recombinant poxviruses FP9 and modified vaccinia virus Ankara. Proc Natl Acad Sci USA 102: 4836-4841. Weisman S, Wang L, Billman-Jacobe H, Nhan DH, Richie TL and Coppel RL. 2001. Antibody responses to infections with strains of Plasmodium falciparum expressing diverse forms of Mz surface protein 2. Infect Immun 69: 959-967. Welch WH. 1897. Malaria: Definitions, Synonyms, history and parasitology. In: Loomis & Thompson's Syst. Pract Med 1: 17. Wipasa J, Xu H, Makobongo M, Gatton M, Stowers A and Good MF. 2002. Nature and specificity of the required protective immune response that develops post challenge in mice vaccinated with the 19-kilodalton fragment of P. yoelii Mz surface protein 1. Infect Immun 70: 6013-6020. Yazdani SS, Shakri AR, Mukherjee P, Baniwal SK and Chitnis CE. 2005. Evaluation of immune responses elicited in mice against a recombinant malaria vaccine based on Plasmodium vivax duffy binding protein. Vaccine 22: 37273737. Journal of Parasitic Diseases: June 2006, Vol. 30, No. 1, 30–36 J PD Isolation and characterization of the paraflagellar rod proteins of Leishmania donovani A. Lahiri and A. Bhattacharya Immunoparasitology Research Unit, Department of Zoology, University of Calcutta, Kolkata. ABSTRACT. The paraflagellar rod (PFR) is a unique cytoskeletal structure present in the kinetoplastid protozoans but absent from their mammalian hosts. It is a massive network of woven cytoskeletal filaments attached to one face of the common 9+2 axoneme of the flagellum. The PFR is necessary for proper parasite motility, viability and successful infection. Biochemical studies carried out on the PFR have revealed that it is composed of both major and minor protein components. In this paper, purification of the PFR proteins of Leishmania donovani has been described, which involves a combination of flagella isolation, non-ionic detergent treatment and restricted proteolysis. Scanning electron microscopy was done to monitor the process of isolation of flagella from intact cell bodies. The major PFR proteins have been identified as two distinct bands of mol wt 76 kDa and 68 kDa, by using sodium dodecyl sulphate polyacrylamide gel electrophoresis analysis. The ultrastructure of the flagellum was studied by using a transmission electron microscope. Keywords: axoneme, flagellum, kinetoplastid, paraflagellar rod INTRODUCTION Leishmania donovani, an intracellular protozoan parasite, causes Kala-azar (visceral leishmaniasis) in humans. The parasite is transmitted by various species of female sandflies (Phlebotomus argentipes). It exists in two morphological forms: the promastigote, residing in the gut of female sandflies, and the amastigote, living in the reticuloendothelial system of the mammalian hosts. The promastigotes possess a full-length, free-flagellum whereas it is rudimentary in amastigotes. The flagellum of promastigotes helps in motility and is involved in hemidesmosomal attachment to the chitinous itima, and maintenance of the parasite within the sandfly gut (Killick-Kendrick, 1979; Killick-Kendrick et al., 1974; Walters et al., 1987). Corresponding author: Dr. A. Bhattacharya, Immuno-parasitology Research Unit, Department of Zoology, University of Calcutta, 35 Ballygunge Circular Road, Kolkata-700 019, W.B., India. The flagellar modes of attachment have been observed with other kinetoplastids also and appear to be essential for their survival within the insect vector (Killick-Kendrick, 1979; Rowton et al., 1981; Vickerman, 1973; Vickerman and Preston, 1976). The flagellum of Leishmania is also involved in chemotactic responses (Bray, 1983). Structurally, the flagellum of Leishmania has a typical 9+2 microtubular axoneme and also possesses a filamentous, lattice-like structure called the paraflagellar rod (PFR) or paraxial rod. The microtubules of the axoneme are arranged in a precise pattern of nine outer double microtubules and two separate central ones. The PFR is a unique cytoskeletal structure found in kinetoplastids, euglenoids and some dinoflagellates (Bastin et al., 1996; Hyams, 1982; Cachon et al., 1988). The paraxial rod of kinetoplastida is similar to that of euglenoids, although apparently, they appear to be morphologically distinct from each other (Farina et 31 The PFR proteins of Leishmania donovani al., 1980; Fuge, 1969; Gallo and Schrevel, 1985). The PFR is a massive network of woven cytoskeletal filaments, running alongside the typical eukaryotic 9+2 axoneme of the flagellum (Russell et al., 1983). Recent molecular studies have demonstrated that the PFR is necessary for proper promastigote motility and viability (Santrich et al., 1997; Bastin et al., 1998). Ablation of a specific molecule of the PFR resulted in mutant cells that were paralyzed, indicating the essential role of PFR in cell motility (Deflorin et al., 1994). Also, ATPase activity was detected in the PFR of euglenoids (Moreira-Leite et al., 1999). The ultrastructure of kinetoplastid PFR appears largely similar in all species of the group (De Souza and Souto-Padron, 1980; Beard et al., 1992). The structure has been divided in three distinct zones in relation to the axoneme namely, proximal, intermediate and distal (Freymüller and Camargo, 1981). The proximal and distal regions each contain filaments of 7-10 nm that intersect at an angle of 100°. The intermediate region contains thin (5 nm) filaments that intersect at an angle of 45° and connect the proximal and distal regions (Maga and LeBowitz, 1999). Attachment filaments extending from axoneme microtubule doublets 4-7 connect the proximal region to the axoneme (Ismach et al., 1989). In Leishmania, the promastigotes possess a full-length flagellum with a PFR, whereas amastigotes contain only an attenuated, non-emergent flagellum completely lacking a PFR. The biochemical composition of the PFR is quite complex and includes both major and minor PFR proteins. Among them, the major PFR proteins have been extensively studied in the parasitic haemoflagellates Trypanosoma cruzi, T. brucei, L. mexicana and L. amazonensis. In these organisms, the major PFR proteins migrate in two bands on sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-PAGE) with mol wt of about 68-75 kDa, and appear to be present in approximately equimolar amounts (Deflorin et al., 1994; Schlaeppi et al., 1989). The major PFR components are conserved in the trypanosomatidae family and form a doublet of homologous proteins in most species of the family (Araujo and Morein, 1991; Saborio et al., 1989). By SDS-PAGE analysis, PFR1 of L. amazonensis migrates at 74 kDa and PFR2 at 69 kDa (Bastin et al., 1996). The PFR1 and PFR2 genes from T. cruzi, T. brucei and L. mexicana are highly conserved across species (over 80% amino acid homology; Maga and Lebowitz, 1999). More complex patterns have been described in T. cruzi in which four major PFR proteins have been identified (Fouts et al., 1998). No significant homology has been reported between major PFR proteins and other known proteins (Imbodem et al., 1995). The ultrastructural study of the flagellum including the PFR and the purification of the PFR proteins provides an important guideline to the researchers for understanding the complete biology of this structure. The parasite L. donovani represents a standard model for studying unique structures such as the PFR because it is one of the most common human pathogens in tropical countries such as India. MATERIALS AND METHODS Parasite culture: The promastigotes were cultured at 22-25° C in liquid M-199 medium supplemented with 10% heat-inactivated fetal bovine serum (FBS, Sigma Aldrich, St. Louis, MO, USA; Morgan et al., 1950). The culture medium was filtered under sterile conditions in a culture room with laminar flow. The medium was kept for 48 h at room temperature to check for any contamination, and then stored at -20° C. Aliquots of the above culture of L. donovani were used in all the experiments. Isolation of flagella: The culture medium containing promastigotes of L. donovani was centrifuged at 2100 x g for 20 min (Cunha et al., 1984). The pellet containing the cells was washed twice in phosphate buffered saline (PBS; 10 mM, pH 7.2; Sigma) and then in buffer A (25 mM TRIS-HCl , 0.2 mM EDTA, 5 mM MgCl2, 12 mM β-mercaptoethanol, 0.32 M sucrose (all from Sigma; pH 7.4) and resuspended in buffer A supplemented with 1% bovine serum albumin (Sigma), 0.1 mM CaCl2 (Sigma), 0.5 mM phenyl methyl sulfonyl fluoride (Sigma) and 5 µg/ml leupeptin (Sigma; Moreira-Leite et al., 1999). The cells were subjected to three different centrifugation speeds: 2600 x g, 3200 x g and 3700 x g, and the flagella were isolated from the cell bodies by centrifugation at an optimum speed of 3200 x g for 30 min. The supernatant containing the flagella was separated from the pelleted and deflagellated cell bodies. This supernatant was then concentrated by centrifugation at 6780 x g for 20 min. The above procedure was carried out at 0-4° C. The pellets containing the flagella were finally suspended in buffer A. Scanning electron microscopy: A scanning electron microscope (SEM) uses a fine beam of electrons to scan back and forth across the metal-coated surface. The principal application of SEM is in the study of 32 surfaces such as those of cells. The aliquots of the treated cells were fixed with 2.5% glutaraldehyde (Sigma) in 0.1M phosphate buffer (pH 7.2-7.4; Sigma) for 24 h and washed in the same buffer for 15 min x 4. The samples were rinsed thrice with double distilled water, each time for 5 min. Then they were dehydrated in different grades of alcohol: 50, 70, 80, 90, 95 and 100% ethanol (20 min each). Gold coating of 200Å was done at 5 mA using Giko Engineering-IB2 ion coater. The cells were dried using vacuum pump and observed under a SEM (model HITACHI S-530), and the photographs were taken by MAMIYA 6 x 7 camera using NOVA 120 ASA films. PFR protein purification: To purify the protein component of PFR, it was necessary to remove the flagellar membrane. The flagellar membranes were removed by non-ionic detergent treatment. Flagellar fractions of L. donovani promastigotes, obtained as described above, were subjected to three rounds of treatment with 2% Nonidet P-40 (Sigma) in PBS (Sigma) at 0-4° C under constant shaking. Each 15 min round of detergent treatment was followed by centrifugation at 17,300 x g for 20 min at 4° C (Moreira-Leite et al., 1999). The pellet of the final centrifugation step was dissolved in PBS (Sigma) and subjected to a brief treatment with 0.0015% trypsin (type XIII, TPCK-treated, Sigma) for 90 s at 28° C. The trypsin treatment was stopped by adding 20-fold excess soyabean trypsin inhibitor (Sigma). The resultant protein fractions were subjected to SDSPAGE. SDS-PAGE: This method is based on the separation of proteins according to mol wt and is particularly useful for monitoring protein purification. The sample to be run on SDS-PAGE was mixed with loading dye (protein:loading dye, 1:1) and boiled for 5 min in a water bath. The stock loading dye had the following composition: double distilled water, 4.8 ml; TRIS (pH 6.8), 1.2 ml; 10% SDS (Sigma), 2 ml; glycerol (Sigma), 1 ml and bromophenol blue (Sigma), 0.5 ml. Before use, 950 µl of stock solution was mixed with 50 µl of β-mercaptoethanol. The sample was run simultaneously with protein markers (Broad Range, Bangalore GENEI, Bangalore, India) in different wells at a constant voltage of 100 V in 10% resolving gel and 4% stacking gel. The gel was then fixed in methanol and stained with Coomassie Brilliant Blue R-250 (Sigma) for a few h, and then washed in destaining solution until clear bands were visible (Laemmli, 1970). Lahiri and Bhattacharya Ultrastructural study of the flagellum by transmission electron microscopy: Cells were fixed in a suitably buffered aldehyde fixative (2.5% glutaraldehyde grade I; Sigma) in 0.1 M sodium cacodylate buffer (pH 7.4; Sigma) at 4° C for 1-4 h. Then the cells were washed for 2 h or overnight at 4° C in three changes of 0.1M sodium cacodylate buffer (pH 7.4). Cells were post-fixed in 1% OsO4 (Sigma) in 0.8% potassium ferricyanide (Sigma) for 1-2 h at room temperature and protected from light (Nakano et al., 2001). The above cells were washed for 5 min x 2 with distilled water. Dehydration was done using the following grades of alcohol: 50, 70, 90 and 95% ethanol, each for 15 min, and 100% ethanol for 15 min x 4. Finally, the treated cells were embedded in Epon polybed 820 epoxy resin. Ultrathin sections were cut and stained with 5% aqueous uranyl acetate (Sigma) and lead citrate (Sigma) and observed under a transmission electron microscope. RESULTS When observed under a SEM, it was revealed that the cells centrifuged at 2600 x g retained their flagella (Fig. 1). The flagella were detached from the intact cell bodies when centrifugation was carried out at 3200 x g (Fig. 2). However, the cells were ruptured when centrifuged at 3700 x g (Fig. 3). SDS-PAGE analysis showed two protein bands of mol wt 76 kDa and 68 kDa that are presumed to be two fractions of the PFR proteins (Fig. 4). Transmission electron microscopy study revealed the ultrastructural details of PFR. The longitudinal section of a flagellum shows that the lattice-like PFR runs parallel to the axoneme even before the emergence of the flagellum from the flagellar pocket 0002 15KV 5um Fig. 1: SEM of a L. donovani cell centrifuged at 2600 x g. 33 The PFR proteins of Leishmania donovani 0016 15KV 5um Fig. 2: SEM of a L. donovani cell centrifuged at 3200 x g. 0021 15KV 5um Flagella Marker Fig. 3: SEM of a L. donovani cell centrifuged at 3700 x g. kD F PFR A BB K 205 97 N 68 29 14 Fig. 4: SDS-PAGE of PFR proteins of L. donovani. The marker proteins are shown on the left side. Fig. 5: Transmission electron microscope photograph of a L. donovani promastigote. [F, flagellum; PFR, paraflagellar rod; A, axoneme; BB, basal body; K, kinetoplast; N, nucleus] (Fig. 5). It also shows the attachment of the flagellum to the basal body at the anterior end of the parasite. The axoneme of the flagellum showed typical eukaryotic 34 9+2 microtubular arrangement. The basal body as well as the base of the flagellum remains ensheathed by a membrane, and the central rod of the flagellum emerges freely. Centrifugation at 2600 x g detached the flagellum partially from the basal body. Complete detachment was obtained at 3200 x g but the flagellum was still surrounded by a membrane. Non-ionic detergent treatment removed the flagellar membranes. DISCUSSION Among protozoans, the kinetoplastid trypanosomes have always held an important position in the scientific research scenario, not only for their medical importance but also for possessing diverse cellular processes. One such unique cytoskeletal structure is PFR. The canonical 9+2 axoneme of the flagellum initiates beating of the latter in most eukaryotes, but the elaborate PFR structure has been observed in only kinetoplastids, euglenoids and some dinoflagellates. Till date, the PFR and its proteins, namely PFR1 and PFR2 have been studied in T. cruzi, T. brucei, L. mexicana and L. amazonensis. These studies revealed that PFR is essential for proper parasite motility and viability, and both these functions are directly attributable to the PFR proteins – PFR1 and PFR2 (Santrich et al., 1997; Bastin et al., 1998). Using new molecular-genetic techniques, PFR1, PFR2 and PFR1/PFR2 null mutants of T. brucei and L. mexicana have been generated. The mutant cells lacked a native PFR structure, showing that despite sharing over 60% amino acid homology and similar physical properties, PFR1 and PFR2 are essential and significantly functional components of PFR. Mutants of L. mexicana having a PFR2-phenotype were unable to colonize in the vector's gut whereas PFR1 and/or PFR2 mutants were viable in axenic culture. This indicates that the PFR plays a key role in the viability of Leishmania in a natural habitat (Hunger-Glaser and Seebeck, 1997). Leishmania parasites lacking PFR were found to display severe disturbances in flagellar waveforms e.g. reduced wavelength and amplitude and decrease in frequency as compared to the beating patterns of normal parasites (Santrich et al., 1997). So, it is of prime importance to isolate and study the ultrastructure of the flagellum and PFR of L. donovani, and to determine its role in the life cycle of the parasite with special reference to PFR proteins. The present studies were carried out with an aim to fulfill the above objectives. Lahiri and Bhattacharya To isolate and purify the PFR proteins, the first essential step was to successfully separate the flagella from intact cell bodies. The process of flagella isolation was monitored by using SEM to make sure that most of the cells remained intact and only lost their flagella. Because the promastigotes were obtained by centrifuging the culture medium at 2000 x g, the cells were subjected to three different higher centrifugation speeds starting from 2600 x g and increasing gradually to 3700 x g to isolate the flagella. When observed under a SEM, it was observed that most of the cells centrifuged at 3700 x g were ruptured (Fig. 3), while those centrifuged at 2600 x g retained their flagella (Fig. 1). Thus, the optimum centrifugation speed for successful isolation of flagella was chosen to be 3200 x g (Fig. 2). The flagellar membranes were removed by treatment with the non-ionic detergent Nonidet P-40. It latter appears to have no effect on the integrity of the flagellar cytoskeleton and is, therefore, unable to break the attachment between the PFR and axoneme (Fig. 5). Beacuse it has been determined that the links between PFR and axoneme are highly sensitive to trypsin (Moreira-Leite et al., 1999), the detachment of PFR from axoneme was accomplished by proteolytic treatment. SDS-PAGE analysis showed two protein bands of mol wt 76 kDa and 68 kDa that are presumed to be two fractions of the PFR proteins (Fig. 4). Extensive studies carried out till date on the major PFR proteins in the parasitic haemoflagellates T. cruzi, T. brucei, L. mexicana and L. amazonensis have shown that the PFR proteins migrate in SDS-PAGE as two bands with mol wt of 70-75 kDa and 68-72 kDa, respectively, and appear to be present in approximately equimolar amounts. So, this result is in conformity with the results reported in previous studies. Transmission electron microscopy study revealed the ultrastructural details of PFR. The longitudinal section of a flagellum shows that the lattice-like PFR runs parallel to the axoneme even before the emergence of the flagellum from the flagellar pocket (Fig. 5). It also shows the attachment of the flagellum to the basal body at the anterior end of the parasite. The axoneme of the flagellum showed typical eukaryotic 9+2 microtubular arrangement (Fig. 5). The basal body as well as the base of the flagellum remains ensheathed by a membrane and the central rod of the flagellum emerges freely. Centrifugation at 2600 x g detached the flagellum partially from the basal body. The PFR proteins of Leishmania donovani Complete detachment was obtained at 3200 x g but the flagellum was still surrounded by a membrane. Nonionic detergent treatment removed the flagellar membranes. The alternation of flagellate and nonflagellate forms of the parasite is attributable to their needs for infection and survival within the host cells. The flagellum is essential for attachment to macrophages, and subsequent penetration and stabilization within macrophages. Apart from attachment, penetration and stabilization within host cells, the main function of flagella is to confer motility and viability to cells so that they can infect host cells. Once the infection process is completed, the presence of flagella is no longer required. So, the flagella become rudimentary and non-functional in the intracellular forms. The present study shows the successful isolation of flagella through scanning electron microscopy. It reports that PFR is composed of two major proteins (mol wt 76 kDa and 68 kDa) in L. donovani, which are unique to that structure and bear no significant homology to other known proteins. Transmission electron microscopy study of the flagellum and PFR provides a relatively complete picture of the biology of these structures. It shows the position of the axoneme and PFR within the flagellum, how they are connected to each other and their structural peculiarities. This study is an important guideline for those who are carrying out researches on the flagellum and PFR of L. donovani. ACKNOWLEDGEMENTS The financial assistance from the Council of Scientific and Industrial Research, New Delhi, [scheme No. 60 (0042)/01/EMR-II] is gratefully acknowledged. REFERENCES Araujo FG and Morein B. 1991. Immunization with Trypanosoma cruzi epimastigote antigens incorporated into iscoms protects mice against lethal challenge in mice. Infect Immun 59: 2909-2914. Bastin P, Mattews KR and Gull K. 1996. The paraflagellar rod of kinetoplastida: solved and unsolved questions. Parasitol Today 12: 302-307. Bastin P, Sherwin T and Gull K. 1998. Paraflagellar rod is vital for trypanosome motility. Nature 391: 548. Beard CA, Saborio JL, Tewari D, Krieglstein KG, Henschen AH and Manning JE. 1992. Evidence for two distinct major protein components, PAR 1 and PAR 2, in the paraflagellar rod of Trypanosoma cruzi. J Biol Chem 267: 21656-21662. 35 Bray RS. 1983. Leishmania: chemotactic responses of promastigotes and macrophages in vitro. J Protozool 30: 322-329. Cachon J, Cachon M, Cosson M-P and Cosson J. 1988. The paraflagellar rod: a structure in search of a function. Biol Cell 63: 169-181. Cunha NL, De Souza W and Hassón-Voloch A. 1984. Isolation and characterization of the paraxial structure of Herpetomonas megaseliae. J Submicrosc Cytol 16: 705713. De Souza W and Souto-Padron T. 1980. The paraxial structure of the flagellum of trypanosomatidae. J Parasitol 66: 229236. Deflorin J, Rudolf M and Seebeck T. 1994. The major components of the paraflagellar rod of Trypanosome brucei are two similar, but distinct proteins which are encoded by two different gene loci. J Biol Chem 269: 28745-28751. Farina M, Attias M and Souto-Padron T. 1980. The paraxial rod of trypanosomatids. J Protozool 33: 552-557. Fouts DL, Stryker GA, Gorski KS, Miller MJ, Nguyen TV, Wrightsman RA and Manning JE. 1998. Evidence for four distinct major protein components in the paraflagellar rod of Trypanosoma cruzi. J Biol Chem 273: 21846-21855. Freymüller E and Camargo EP. 1981. Ultrastructural differences between species of trypanosomatids with and without endosymbionts. J Protozool 28: 175-182. Fuge H. 1969. Electron microscopic studies and the intraflagellar structure in trypanosomes. J Protozool 16: 460-465. Gallo JM and Schrevel J. 1985. Homologies between paraflagellar rod proteins from trypanosomes and euglenoids revealed by a monoclonal antibody. Eur J Cell Biol 36: 163-168. Hunger-Glaser I and Seebeck T. 1997. Deletion of the genes for the paraglagellar rod protein PFR-A in Trypanosoma breecei is probably lethal. Mol Biochem Parasitol 90: 347351. Hyams J. 1982. The Euglena paraflagellar rod: structure, relationship to other flagellar components and preliminary biochemical characterization. J Cell Sci 55: 199-210. Imbodem M, Müller N, Hemphill A, Matioli R and Seebeck T. 1995. Repetitive proteins from the flagellar cytoskeleton of African trypanosomes are diagnostically useful antigens. Parasitol 110: 249-258. Ismach R, Ciani CML, Caulfield JP, Langer PJ, Hein A and McMahon-Pratt D. 1989. Flagellar membrane and paraxial rod proteins of Leishmania: characterization employing monoclonal antibodies. J Protozool 36: 617-624. Killick-Kendrick R. 1979. Biology of Leishmania in phlebotomine sandflies. In: Biology of the Kinetoplastida. Lumsden WHR and Evans DA (Eds) Academic Press, New York. pp. 203-260. 36 Killick-Kendrick R, Molyneux DH and Ashford RW. 1974. Leishmania in phlebotomine sandflies I. Modification of the flagellum associated with attachment to the midgut and oesophageal valve of the sandfly. Proc R Soc Lond B. 187: 409-419. Laemmli UK. 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 277: 680685. Lahiri and Bhattacharya paraflagellar rod of Crithidia fasciculata. Eur J Cell Biol 30: 137-143. Saborio JL, Hernandez JM, Narayanswami S, Wrightsman R, Palmer E and Manning JE. 1989. Isolation and characterization of paraflagellar proteins from Trypanosoma cruzi. J Biol Chem 264: 4071-4075. Maga JA and LeBowitz JD. 1999. Unraveling the kinetoplastid paraflagellar rod. Trends in Cell Biol 9: 409-413. Santrich C, Moore L, Sherwin T, Bastin P, Brokaw C and Gull K. 1997. A motility function for the paraflagellar rod of Leishmania parasites revealed by PFR-2 gene knockouts. Mol Biochem Parasitol 90: 95-109. Moreira-Leite FF, De Souza W and Cunha-e-Silva NL. 1999. Purification of the paraflagellar rod of trypanosomatid Herpetomonas megaseliae and identification of some of its minor components. Mol Biochem Parasitol 104: 131-140. Schlaeppi K, Deflorin J and Seebeck T. 1989. The major component of the paraflagellar rod of Trypanosoma brucei is a helical protein that is encoded by two identical, tandemly linked genes. J Cell Biol 109: 1695-1709. Morgan JF, Mortan H and Parker RC. 1950. Nutrition of animal cells in tissue culture. I. Initial studies on a synthetic medium. Proc Soc Exp Biol Med 73: 1. Vickerman K. 1973. The mode of attachment of Trypanosoma vivax in the proboscis of the Tsetse fly Glossina fuscipes and ultrastructure of the epimastigote stage of the trypanosome. J Protozool 20: 394-404. Nakano MAY, Nagakura K and Aikawa M. 2001. Invasive forms of Toxoplasma gondii, Leishmania amazonensis and Trypanosoma cruzi have a positive charge at their contact site with host cell. Parasitol Res 87: 193-197. Rowton ED, Lushbaugh WB and McGee RB. 1981. Ultrastructure of the flagellar apparatus and attachment of Herpetomonas ampelophilae in the gut and malpighian tubules of Drosophila melanogaster. J Protozool 32: 297301. Russell DG, Newsam RJ, Palmer GCN and Gull K. 1983. Structural and biochemical characterization of the Vickerman K and Preston TM. 1976. Comparative study of kinetoplastida flagellates. In: Biology of the Kinetoplastida. Lumsden WHR and Evans DA (Eds) Academic Press, New York. pp 11-34. Walters LL, Modi GB, Tesh RB and Burrage T. 1987. Hostparasite relationship of Leishmania mexicana mexicana and Lutzomyia abennenci (Diptera: Psychodidae). Am J Trop Med Hyg 36: 294-314. Journal of Parasitic Diseases: June 2006, Vol. 30, No. 1, 37–40 J PD Onchocercosis in Benue state, Nigeria: comparative epidemiological studies amongst the Etulo and Idoma ethnic groups E. A. Omudu and B. O. Atu Department of Biological Sciences, Benue State University, Makurdi. ABSTRACT. The prevalence of onchocercosis caused by Onchocerca volvulus amongst two distinct ethnic groups in Benue State was investigated using Rapid Assessment Methods (RAMs) and skin snip technique. Results showed that of the 2126 individuals examined in the Idoma-speaking Okpokwu Local Government Area, 16.6% were positive either for Leopard skin (LS) or palpable nodules. While of the 1005 individuals examined in the Etulo area, 5.6% had either LS or palpable nodules. The prevalence rate varied significantly (p < 0.05). Correlation analysis showed significant association between LS, palpable nodules and aging (r=0.43, p < 0.001). Skin snip results showed communities with higher LS and nodules had higher community microfilarial load. Correlation coefficient r=0.64 was obtained when skin snip prevalence was compared with RAMs in the Etulo area. The implications of these results were discussed especially in terms of prioritizing the implementation of community-directed treatment with ivermectin. Keywords: microfilariae, Nigeria, onchocercosis, Rapid Assessment Methods INTRODUCTION Onchocercosis (river blindness) is a parasitic disease caused by the nematode Onchocerca volvulus and transmitted by the bites of blackflies belonging to Simulium damnosum complex. It is estimated that over 81 million people are at risk of infection, some 18 million already infected and one million people visually impaired of whom about 340,000 are blind (WHO 1995). Apart from ophthalmological degeneration leading to blindness, the disease also presents bizarre dermatological, lymphatic and systemic manifestations (Edungbola et al., 1990). Onchocercosis has been reported in some parts of Benue State, (Gemade and Dipeolu, 1983; Amuta and Olusi, 2000), where it is affecting socio-economy of Corresponding author: Dr. Edward Agbo Omudu, Department of Biological Sciences, Benue State University, P. M. B. 102119, Makurdi, Nigeria. E-mail: eddieomudu@yahoo. com human population. The development and use of Rapid Assessment Methods (RAMs) using prevalence of Leopard skin (LS) and palpable nodules for diagnosis of onchocercosis and free distribution of ivermectin for treatment has heightened expectation that the disease can be contained. For this to happen, there is a need to know the extent of the disease especially in communities without baseline information. This study, using RAM, was undertaken to provide pretreatment prevalence and identify communities eligible for mass distribution of ivermectin. MATERIALS AND METHODS Study areas: The study areas comprised of nineteen Idoma speaking villages in Okpokwu Local Government Area (LGA) and twelve Etulo speaking villages in Buruku and Katsina-Ala LGA. There are distinct cultural and occupational differences that may influence exposure to biting blackflies and perception of disease manifestation. The vegetation and 38 topographical features of the study areas have been described by Nwoke et al. (1998) as being favourable and suitable for onchocercosis transmission. Okpokwu: The local population of this area (8°00 and 8° 30 E and 6° 58 and 7° 25 N) practice peasant agriculture, fishing and hunting are secondary occupations. With numerous breeding sites on the Okpokwu river system that drain these villages and outdoor activities performed by the population, they are continually exposed to bites of the vectors. Buruku and Katsina-Ala: The Etulo community inhabits twelve villages which are bisected by river Katsina-Ala, a major tributary of Benue River. Majority of the villages are located along the bank of the river with fishing and farming constituting the major occupation. Apart from river Katsina-Ala, another perennial river known as Ogaturu drains most of Etulo land. The two predominant rivers are fast flowing and harbour larvae and pupae of S. damnosum complex (Nwoke et al., 1998; Gemade and Dipeolu, 1983). RAMs for diagnosis: Based on the analysis of large volumes of epidemiological data on onchocercosis, depigmentation of skin LS and palpable onchocercal nodules were recommended as rapid assessment procedures to determine communities eligible for treatment with ivermectin. The merits and limitations of these methodologies are summarized by Edungbola et al. (1993). Community: Only adults 18 years and above were examined for presence of nodules and LS. All persons sampled were engaged in one or more rural occupation and have resided in the community for at least five years (Gemade, 1993). Communities were visited a day prior to the fixed date for examination in order to mobilize people for the survey and free treatment. Search for nodules: For search of nodules, the patients were requested to strip down to the loin. As the person stretched his arms above the head, the whole chest region was examined both visually and by palpation. Special attention was given to iliac crest around knees, ankles, ribs (from and rear), chest wall, shoulders, elbows, wrist and head. Search for LS: The search for LS on patients was more specific in location and faster as typical onchocercal depigmentation commonly occurs on lower limb of adults above 30 years. Patients were asked to pull up trousers and/or wrappers to the thigh region. Skin snip examination: Skin snip parasitological procedure for identification of microfilariae of O. Edward and Bernard volvulus was carried out in whole population of the Etulo area; however, this was not possible in Okpokwu LGA as a result of immunological and parasitological studies just concluded in the same area by Amuta and Olusi (2000). Two snips, one each from the right and left iliac crest were taken from each individual with the aid of a 2 mm bite corneoscleral punch (E-2802, Holt Storz, Germany). Each snip was incubated in a well of microtitre plate containing normal saline. The plates were then examined within 30 min to one hour by microscopy for the presence of O. volvulus microfilariae (Wentworth, 1988). RESULTS AND DISCUSSION Out of 31 villages examined, all were found to have individuals with LS, whereas 27 (87%) villages were carrying nodules of onchocercal origin. Out of 2126 individuals examined from the Idoma speaking villages in Okpokwu LGA, 248 (11.6%) had LS, 105 (4.9%) had nodules and 35 (1.6 %) had both nodules and LS while 353 (16.6%) had either LS or nodules (Table 1). In 1005 Etulo-speaking individuals, 45 (4.5%) had LS, 12 (1.2%) had nodules and 2 (0.2 %) had both LS and nodules while 57 (5.6%) had either LS or nodules (Table 2). The number of cases with LS, visible or palpable nodules observed in the Okpokwu area varied significantly (p < 0.05) from that of Etulo area. Whereas LS in Okpokwu area ranged from 7.3–28.8%, in Etulo area it ranged from 0.8–14.0%. Only one village (Ashitenaku) had 14% LS, whereas all the other Etulo villages had LS below 7.3%. In Okpokwu, the prevalence of onchocercal nodules ranged from 2.2%–10.9% and in Etulo area it ranged from 0.8–3.1% only one village (Ugye) in Etulo had nodule prevalence of 3.1%, whereas all other villages had the prevalence below 2.3%. LS and onchocercal nodules were mostly noticed in individuals within 40–69 years of age, highest (48.7%) being in between 50–59 years. This same age group accounted for 20.5% of onchocercal nodules. Correlation analysis to determine association between these onchocercal manifestations with age showed significant correlation (r = 0.45, p < 0.001). The patients complained varying degrees of visual impairment but, only five cases of total blindness were recorded. One classical blind case was seen in a 61 years old man in Effa (Okpokwu), who had Leopard skin on both shins and a nodule on the knee. The Etulo area on the other hand had four cases of blindness (two in Ashitenaku, one each in Oglazi and Agbou respectively). Skin snip examination in the Etulo area showed 536 (53.3%) persons were positive for O. volvulus microfilariae. The prevalence by villages varied with 39 Epidemiology of onchocercosis in Nigeria Table I: Determination of the prevalence of onchocercosis in Okpokwu villages by using LS and palpable nodules as diagnostic methods Villages No. of persons examined No. positive LS (%) No. positive for nodules (%) No. positive for both nodules and LS (%) Ojoga Ai-Ebiega Ipole Iwewe Obotu Opialu Ollo Ai-Ohida Ai-Okpe Ogene Ijege Ogege Effa Idobe Oto-Oklenyi Odokpo Ai-Akpa Aidogodo Ede 82 51 49 45 85 132 72 72 80 102 90 250 92 60 52 68 58 418 268 23(28.8) 11(21.5) 10(20.4) 3(6.6) 12(14.1) 13(9.8) 8(11.1) 6(8.3) 10(12.5) 15(14.7) 9(10.0) 22(14.6) 21(14.6) 6(10.0) 6(11.5) 5(7.3) 10(17.2) 36(8.6) 22(8.2) 9(10.9) 3(5.8) 2(4.0) 2(4.4) 8(9.4) 10(7.5) 5(6.9) 2(2.7) 3(3.7) 3(2.9) 2(2.2) 8(3.2) 10(10.8) 6(10.0) 3(5.7) 2(2.9) 2(3.8) 15(3.5) 10(3.7) 5 ( 6.1 ) 2 ( 3.9 ) 3 ( 3.5 ) 4 ( 3.0 ) 1 (1.3 ) 1 (1.3 ) 1 (0.9 ) 1 (0.4 ) 5 (5.4 ) 2 (3.3 ) 1 (1.4 ) 3 (5.1 ) 4 (0.9 ) 2 (0.7 ) Total 2126 248 (11.6) 105 (4.9) 35 (1.6) Table II: Determination of the prevalence of onchocercosis in Etulo villages by using LS, palpable nodules and skin snip as diagnostic methods Villages No. of persons examined No. of positive LS (%) No. of positive nodules (%) No. positive for both LS and nodules (%) No. of positive skin snip (%) Agbou Agbatala Agia Angwauje Ashitenaku Oglazi Otsaazi Otsafu Ugye Ogurube Otanga Okpashila 64 55 87 92 100 68 99 47 95 94 84 120 2(3.1) 2(3.6) 1(1.1) 2(2.1) 14(14.0) 5(7.3) 9(9.0) 3(6.3) 2(2.1) 2(2.1) 2(2.3) 1(0.8) 1(1.8) 2(2.3) 1(1.0) 1(1.4) 2(2.0) 1(2.1) 3(3.1) 1(0.8) 1(1.0) 1(1.05) - 40(62.5) 44(80.0) 39(44.8) 44(47.8) 74(74.0) 46(67.6) 59(59.5) 31(65.9) 31(34.0) 38(40.4) 29(34.5) 61(50.8) Total 1005 45 (4.5) 12 (1.2) 2 (0.2) 536 (53.3) 40 four villages being meso-endemic (60–89%) while the other eight were hypo-endemic (< 60%; Table 2). Communities with higher prevalence of LS and onchocercal nodule also have proportionately higher prevalence of skin microfilariae. Correlation coefficient (r) = 0.64 was obtained when skin snip prevalence was compared with RAMs; there was, however, no correlation between microfilarial prevalence and visual impairment (r) = 0.017 in the Etulo area. The prevalence of onchocercosis varied significantly (p < 0.05) in the different villages. Others have also reported varying endemicity within same biogeographical zones (Nwoke et al., 1994; Nocks et al., 1998). It may be due to differences in duration and degree of exposure of members from different communities to bites of infected vectors (Nwoke et al., 1994) and immunological factors (Murdock, 1992). Another reason might be topographical differences, whereas Okpokwu area is characterized by hilly terrain with many fast flowing streams, the Etulo area is predominantly flat plain with few fast flowing streams. A total of 479 (89.4%) individuals who had microfilariae in their skin biopsy showed no clinical sign of nodules or LS. Many other studies in Nigeria where skin snip, LS and palpable nodules were combined to determine onchocercosis prevalence revealed similar findings where LS and visible nodules varied significantly (p < 0.05) with the community microfilarial rate (Nwoke et al., 1994; Nock et al., 1998). Amuta and Olusi (2000) also reported high prevalence rate using skin snip in some villages where we recorded lower prevalence using LS and onchocercal nodules. The very low incidence of blindness recorded in both study sites agreed with Nwoke et al. (1994) and has a common observation because rainforest region of Nigeria is endemic for O. volvulus strains that rarely cause blindness unlike to savanna types which are more invasive and more pathogenic to the eyes (Nwoke et al., 1994). The use of LS and palpable nodules as diagnostic procedures for onchocercosis are highly desirable as they are cheap, fast, convenient and highly efficient (Carme et al., 1993; Edungbola et al., 1993; Nwoke et al., 1998; Nock et al., 1998). A combination of these two methods as done in this study is of exceptional advantage since it identifies community's endemicity. There is a need to commence treatment with ivermectin in some of the villages investigated, especially those with LS prevalence of more than Edward and Bernard 10%. It is also important to train communities on how to conduct rapid assessment procedures in order to prioritize community eligibility. ACKNOWLEDGEMENTS We acknowledge with thanks the assistance and support of National Onchocercosis Control Programme Coordinators for Buruku, Katsina-Ala and Okpokwu LGAs in carrying out the present studies. REFERENCES Amuta EO and Olusi TA. 2000. Sero-epidemiological study of Onchocerca volvulus using eluate of blood collected on filter paper. Nigerian Journal of Parasitology 21:33-38. Carme B, Samba Y, Ntsoumou MV and Yebakima A. 1993. Prevalence of depigmentation of skin: a simple and cheap way to screen for severe endemic onchocercosis in Africa. Bulletin of the World Health Organization 70: 755-758. Edungbola LD, Babata AL, Asaolu SO, Duke BOI and Connor DH. 1990. Leopard skin and onchocercosis. Nigerian Journal of Parasitology 9:77-82. Edungbola LD, Nwoke BEB, Onwurili COE, Akpa AUC and Tayo-Mafe M. 1993. Selection of rapid assessment methods for community diagnosis of onchocercosis. Nigerian Journal of Parasitology 12:45-50. Gemade EI and Dipeolu OO. 1983. Onchocerciasis in Benue State of Nigeria. II. Prevalence of the disease among the Tivs living in the Kwande local government area. Annals of Tropical Medicine and Parasitology 77:513-516. Gemade EI. 1993. Training manual in Rapid Assessment Methods using nodules and Leopard skin, treatment schedule based on weight and height and health education for large-scale distribution of Mectizan. Revised and updated by sight savers international (Nigeria). pp 42. Murdock ME. 1992. The skin and the immune response in onchocercosis. Tropical Doctor (Supplementary) 1:44-62. Nock IH, Ripiye P and Galadima M. 1998. Diagnostic value of nodules and Leopard skin in community assessment of human onchocercal endemicity. Nigerian Journal of Parasitology 19:19-24. Nwoke BEB, Edungbola LD, Mencias BS, Njoku AJ, Abanob OC, Nwogu, FU et al. 1994. Human onchocercosis in rain forest zone of southern-eastern Nigeria. 1: Rapid assessment methods for community diagnosis in Imo river basin. Nigerian Journal of Parasitology 15:7-18. Nwoke BEB, Dozie INS, Gemade EI and Jiya JY. 1998. The present status of human onchocercosis in southeastern Nigeria using rapid epidemiological mapping (REMO). Nigerian Journal of Parasitology 19:11-18. Wentworth BB. 1988. Diagnostic Procedures for Mitotic and parasitic infections. American Public Health Association Publication Inc. Washington DC. 637 pp. WHO. 1995. Expert Committee Report on Onchocercosis. Fourth Report, Technical Report Series. 253 pp. Geneva. Journal of Parasitic Diseases: June 2006, Vol. 30, No. 1, 41–44 J PD Distribution of iron in plasma, erythrocytes and tissues of calves with the progression of Theileria annulata infection 1 N. Sangwan and A. K. Sangwan 1 2 2 Department of Veterinary Biochemistry, CCS Haryana Agricultural University, Hisar. Department of Veterinary Parasitology, CCS Haryana Agricultural University, Hisar. ABSTRACT. The distribution of iron in plasma, erythrocytes and tissues was studied in relation to the progression of Theileria annulata infection in cross-bred, 4-6 months old male calves. Group-I animals were experimentally infected with Theileria annulata by attaching ten Theileria positive ticks on the ear of each calf and Group-II was kept as a healthy uninfected control. The blood and plasma samples were collected at 0, 10, 15, 20 and 25 days post-infection. The tissue samples of liver and spleen were collected immediately after the death of the infected calves. The iron concentrations were estimated by using atomic absorption spectrophotometer. The infected calves had significantly (p < 0.05) low levels of iron in whole blood and erythrocytes, whereas the plasma iron levels did not change much. The liver and spleen iron concentrations increased significantly (p < 0.05) as compared to the normal range reported in the literature. This study suggests that care should be taken while treating theileriosis cases by not supplementing with iron, but by administering agents which minimize oxidative damage and help in the better utilization of iron for erythropoiesis. Keywords: cattle, haematology, iron, liver, spleen, Theileria annulata INTRODUCTION Tropical theileriosis is a tick-borne disease of cattle, which causes morbidity and loss of productivity in zebu cattle and lethal disease in European and crossbred stock (Gill et al., 1977; Purnell, 1978). Theileriosis is characterized by fever and lymphoproliferative disorders, which may be associated with leucopenia, wasting, anaemia and jaundice in susceptible cattle (Preston et al., 1992). The damage to the host is caused by both schizonts in lymphocytes/monocytes and piroplasms in erythrocytes, resulting in lymphadenopathy and Corresponding author: Dr. Nirmal Sangwan, Department of Veterinary Biochemistry, CCS Haryana Agricultural University Hisar-125 004, Haryana, India. E-mail:ns@hau.ernet.in haemolytic anaemia with icterus. Serum iron has been found to be reduced significantly in Friesian cattle naturally infected with Theileria annulata (Omer et al., 2003), whereas, on the contrary, it increased in Holstein calves infected with T. sergenti; concentrations of non-haem iron and ferritin in liver and spleen were also significantly higher in infected calves (Watanabe et al., 1998). Therefore, to know whether the animals suffering from theileriosis should be supplemented with iron or not, the present studies were conducted to estimate the status of iron in blood, plasma, erythrocytes and tissues of T. annulatainfected calves. MATERIAL AND METHODS Two groups of six cross-bred, 4–6 months old male calves were de-wormed, sprayed with acaricides and 42 vaccinated against Foot and Mouth Disease. The animals were fed concentrate as per National Research Council (1988). Wheat bhoosa (roughage) and green fodder were provided ad libitum. After 15 days, Group-I animals were experimentally infected with T. annulata by attaching ten Theileria positive ticks (five pairs of females and males) on the ear of each calf, and the other group was kept as a healthy uninfected control. Infection was monitored by daily clinical examination (rectal temperature and lymph node palpation) and at 2-day intervals, Giemsa-stained thin blood and lymph node biopsy smears were examined after the appearance of lymphadenopathy. The day of the death of infected calves was recorded. The blood and plasma samples were collected in heparinised and sterilized glass tubes on 0, 10, 15, 20 and 25 days postinfection. The tissue samples of liver and spleen were collected immediately after the death of infected calves (four animals). The iron concentrations in different samples were estimated by using atomic absorption spectrophotometery (Dunkan, 1976), following acid digestion of organic matter and by using calibration standards made in 0.1N HCl. The samples of blood (1 ml) and plasma ( 2 ml) were digested separately with 10 ml of digest acid (4:1; nitric:perchloric acid ), dried, washed x 2 with 5 ml of deionised water, dried each time and reconstituted with 10 ml and 5 ml of 0.1 N HCl, respectively. The tissue samples of liver and spleen ( 0.25 g , dried and ground ) were also digested as for plasma and then reconstituted in 10 ml of 0.1 N HCl. Simultaneously, blanks were also run. The iron concentrations in erythrocytes were calculated by using the following formula: trace-element in erythrocytes/ml of blood = whole blood-plasma (1-packed cell volume/100). Haemoglobin and haematocrit were estimated by using cyanmethaemoglobin and microhaematocrit methods, respectively (Schalm et al., 1975). The data were subjected to standard error of means (SE) and Analysis of Variance (ANOVA) for statistical significance (Snedecor and Cochran, 1967). RESULTS AND DISCUSSION During the course of Theileria infection, the calves showed fever, anaemia, anorexia, cachexia, diarrhoea, respiratory distress and recumbancy. The clinical and parasitological findings are recorded in Table I. The death of infected calves occurred between 18–27 days post-infection. The haematological responses of calves are given in Table II. As the disease progressed, marked fall in hemoglobin and packed cell volume (PCV) were observed. The values for hemoglobin and PCV were reduced significantly (p < 0.05) from the Sangwan and Sangwan initial mean value of 9.88 ± 0.14 (g%) and 30.71 ± 0.63 (%) to 2.9 ± 0.5 (g%) and 9.0 ± 1.41(%), respectively, on day 25 post-infection. The clinical and haematological findings of bovine tropical theileriosis reported here resembled those documented by other authors (Preston et al., 1992; Sahu et al., 1996; Forsyth et al., 1999). The infected calves had significantly (p < 0.05) low levels of iron in whole blood and erythrocytes, whereas in plasma, the levels of iron did not change much (Table II). The levels of iron in blood and erythrocytes reduced significantly (p < 0.05) from 266.9 ± 7.38 to 67.9 ± 8.37 µg/ml and 384.3 ± 12.46 to 72.7 ± 10.7 µg/ml, respectively, on day 25 postinfection. Generally, the risk of iron inadequacy is assessed by measuring blood hemoglobin and/or the highly correlated PCV. However, in the study under report, the low levels of hemoglobin and PCV did not correspond to the low levels of iron in the body reserves. The liver and spleen iron concentrations were found to be 628.8 ± 78.23 [µg/g dry matter (DM)] and 1647.7 ± 182.53 (µg/g DM), respectively. These values in infected group are much higher than the range reported by Georgievskii et al., 1982 i.e. 180–376 µg/g DM for liver of adult cattle and 200–400 µg/g on fresh basis for spleen, respectively (Georgievskii, 1982). Therefore, it is important to distinguish anaemia associated with Theileria infection from that caused by iron deficiency. Iron deficiency is indicated by low levels of iron in the liver, and a marginal band of 150–250 µg/g DM is tentatively proposed to separate deficient from normal claves (Green et al., 1993). The high levels in liver and spleen could be attributed either to the increased haemolysis or animals' limited capacity to excrete iron (Kreutzer and Kirchgessner, 1991). If there would have been haemolysis that would have resulted in high plasma iron concentration. But in the present study, the plasma iron levels did not increase, which point towards the elimination of infected erythrocytes by phagocytosis. Also, while studying the pathogenesis of anaemia in T. annulata infection, Hooshmand-Rad (1976) suggested that an autoimmune reaction was largely responsible for the development of anaemia and postulated that the production of antibodies was triggered by the development of schizonts; erythrocytic forms apparently were not involved. This contention has been supported by the finding that autohaemagglutinin antibodies were detected only in cases of theileriosis due to a field or an agamogenous strain (lacking erythrocytic forms) but not in premune 43 Iron distribution in Theileriosis Table I. Host responses (clinical and parasitological) in calves infected with Theileria annulata Clinical observations Parasitological observations Thermal reaction Group Calf No. Maximum (°F) Appearance (day) Piroplasms Max. Appearance (%) (day) (died on day) Max. (%) 1 2 3 4 5 6 4 5 4 4 5 4 9 9 9 8 9 8 105.5 104.8 105.8 106.0 104.4 106.0 10 10 10 9 10 10 15 10 20 10 25 10 13 13 13 11 13 13 85 14 25 20 15 20 7-12 - - - - - - - I II Lymph- Commencement adenopathy (day) (day) Macroscizonts Result 23rd 29th 27th 18th 19th 20th - Group-I: calves were experimentally infected with Theileria annulata by attaching ten positive ticks on the ear of each calf. Group-II:healthy uninfected control. Table II. Estimated values for haemoglobin, packed cell volume and iron in the blood, plasma and erythrocytes of cross-bred calves infected with Theileria annulata1 Parameters Days post-infection 0 10 15 20 25 Hb (g/dl) 9.88a ± 0.14 6.03b ± 0.48 5.88b ± 0.41 5.10c ± 0.42 2.90d ± 0.5 PCV (%) 30.71a ± 0.63 21.42b ± 0.69 20.60b ± 0.36 20.00b ± 1.88 9.00c ± 1.41 Blood (µg/ml) 266.9a ± 7.38 188.5b ± 12.7 165.9c ± 10.31 151.1cd ± 11.77 67.9e ± 8.37 Plasma (µg/ml) 2.00a ± 0.05 1.52ab ± 0.21 1.33b ±0.18 1.64ab ± 0.05 2.03ab ± 0.35 Erythrocytes (µg/ml of blood) 265.5a ± 7.39 187.3b ± 12.81 164.8cb ± 10.27 149.8c ± 11.83 66.1d ± 8.72 1 Calves were experimentally infected with Theileria annulata by attaching ten positive ticks on the ear of each calf. Hb, hemoglobin; PCV, packed cell volume. 2 Values with common superscripts do not differ significantly (p < 0.05). splenectomised calves. Only mild anaemia and bilirubinaemia occurred in the premune splenectomised calves in spite of the high level of parasite load and it was suggested that the infected erythrocytes may be eliminated by augmented phagocytosis. During infection, iron is redistributed by the host in an attempt to deplete the pathogen of iron (Weinberg, 1984). The redistribution of iron by the host may cause the secondary anaemia by depriving iron to erythropoietic tissues. At a time when the liver and spleen iron levels are already high, additional iron supplementation in the Theileria-infected animals could result in increase in the liver iron concentration to toxic levels. The liver iron concentration of 1000 µg/g DM is considered hepatotoxic (Underwood and Suttle, 1999) and sufficient reactive iron may be available to cause peroxidative damage to liver (Kent and Bahu, 1979), and this may be the underlying 44 pathogenic mechanism of peroxidative damage to lipid membranes (Gordeuk et al., 1987). The extent of injury depends on the antioxidant status of the animal, particularly its vitamin E status (Omara and Blakeley, 1993; Ibrahim et al., 1997). Caeruloplasmin, a copper containing protein, has also been reported to provide antioxidant defences by scavenging free-iron and free-radicals (Saneko et al., 1994). In summation, clinical signs in terms of anaemia, low levels of iron in whole blood and erythrocytes and the presence of high concentrations of iron in liver and spleen suggest that T. annulata can harm its host by disrupting the normal functioning of liver and spleen, upon which effective protective immune responses and biochemical mechanisms depend. So, it is important not to supplement Theileria-infected animals with iron, but rather give them other nutrients which help in preventing the oxidative damage (such as vitamin E) and better utilization of iron for erythropoiesis. ACKNOWLEDGEMENTS This work was carried out at CCS Haryana Agricultural University, Regional Research Station, Uchani, Karnal, and was financially supported by the National Agricultural Research Project of the Indian Council of Agricultural Research. REFERENCES Dunkan L (Edt.).1976. Chemical Analysis by Atomic Absorption Spectrophotoscopy. (Third Ed). Lea and Febiger, Philadelphia. Forsyth LMG, Minns FC, Kirvar E, Adamson RE, Hall FR, McOrist S, Brown CGD and Preston PM.1999. Tissue damage in cattle infected with Theileria annulata accompanied by metastasis of cytokine-producing, schizont infected mononuclear phagocytes. Journal of Comparative Pathology. 120:39-57. Georgievskii VI, Annenkov BN and Samokin VI (Eds.).1982. Studies in the Agricultural and Food Sciences. Mineral Nutrition of Animals. Butterworths, London, pp 171-224. Gill BS, Bhattacharyulu Y and Kaur D.1997. Symptoms and pathology of experimental bovine tropical theileriosis (Theileria annulata infection). Annales de Parasitologie. 52:597-608. Gordeuk VR, Bacon BR and Brittenham GM. 1987. Iron overload: Cause and consequences. Annual Review of Nutrition. 7:485-508. Green LE, Berriatua E and Morgan KL.1993. Anaemia in housed lambs. Research in Veterinary Science. 54:306-311. Ibrahim W, Lee V-S, Ye C-C, Szabo J, Bruckner G and Chow CK. 1997. Oxidative stress and antioxidant status in mouse liver: effects of dietary lipid, vitamin E and iron. Journal of Nutrition. 127:1401-1406. Sangwan and Sangwan Hooshmand-Rad P.1976. The pathogenesis of anaemia in Theileria annulata infection. Research in Veterinary Science. 20:324-329. Kent G and Bahu RM. 1979. Iron overload. In: MacSween RNM, Anthony PP and Schewr PJ (Eds). Pathology of the liver. Churchill Livingstone, Edinburgh, pp 148-163. Kreutzer M and Kirchgessner M.1991. Endogenous iron excretion: a quantitative means to control iron metabolism. Biology of Trace Element Research. 29:77-92. Naessens J, Grab DJ, Fritisch G.1996. Characterization of bovine transferring receptor on normal activated and Thieleria parva transformed lymphocytes by a new monoclonal antibody. Veterinary Immunology and Immunopathology. 52:65-76. National Research Council.1988. Nutrient Requirements of Dairy Cattle. Sixth Ed. National Academy of Science. Washington, D.C., USA. Omara FO and Blakeley BR. 1993. Vitamin E is protective against iron toxicity and iron-induced hepatic vitamin E depletion in mice. Journal of Nutrition. 123:1649-1655. Omer OH, El-Malik KH, Magzoub M, Mahmoud OM, Haroun EM, Hawas A and Omar HM. 2003. Biochemical profiles in Friesian cattle naturally infected with Theileria annulata in Saudi Arabia. Veterinary Research Communication. 27:15-25. Preston PM, Brown CGD, Bell-Sakyi LJ, Richardson W and Sanderson A. 1992. Tropical theileriosis in Bos Taurus and Bos indicus calves. Response to infection with graded doses of sporozoites. Research in Veterinary Science. 53:230-243. Purnell RE. 1978. Theileria annulata as a hazard to cattle in the countries on the northern Mediterranean littoral. Veterinary Research Communication. 2:3-10. Sahu PK, Misra SC, Panda DN and Mohapta M. 1996. Haematological and biochemical alterations in Theileria annulata infected cross-bred cattle. Indian Veterinary Journal. 73: 995-997. Saneko EL, Yaroplov AI and Harris ED. 1994. Biological functions of caeruloplasmin expressed through copperbinding sites. Journal of Trace Elements and Experimental Medicine. 7: 69-88. Schalm OW, Jain NC and Caroll EJ (Eds.). 1975. Veterinary Haematology. Third Ed., Lea and Fabiger, Philadelphia. Snedecor GW and Cochran WJ.1967. Statistical Methods. Seventh Ed. Oxford and IBG Publishing Co., New Delhi. Underwood EJ and Suttle NF. 1999. The Mineral Nutrition of Livestock. Third Ed., CAB International Publishing Co. p 361. Watanabe K, Ozawa M, Ochiai H, Kamohara H, Lijima N, Negita H, Orino K and Yamamoto S. 1998. Changes in iron and ferritin in anaemic calves infected with Theileria sergenti . Journal of Veterinary Medical Science. 60:943947. Weinberg ED. 1984. Iron withholding: a defence against infection and neoplasia. Physiological Reviews. 64:65-102. Journal of Parasitic Diseases: June 2006, Vol. 30, No. 1, 45–52 J PD Mosquito breeding in riceland agro-ecosystem near Chennai, Tamil Nadu, India 1 J. Ravindran and J. Williams 2 1 Integrated Disease Vector Control Project (National Malaria Research Institute), Field Station, Chennai. P. G. and Research Department of Zoology, Loyola College, Chennai. 2 ABSTRACT. Breeding ecology of mosquitoes was studied in riceland area near Chennai during the period 1992-1999. The study area was a transition zone with withdrawal of intense agricultural activities to facilitate urbanization. A myriad of mosquitoes including potential vectors of malaria and Japanese encephalitis (JE) were observed breeding in various habitats present. A longitudinal survey showed no malaria vector breeding in rice fields. Extensive breeding of Culex tritaeniorhynchus (JE vector) was observed. Natural breeding pattern indicated peak anopheline and culicine density during first two weeks after transplantation. Pre- and post-monsoon surveys in irrigation wells indicated breeding of 16 species of mosquitoes. Both Anopheles stephensi and An. culicifacies, the urban and rural malaria vectors, respectively, were encountered breeding in the wells; An. stephensi breeding was predominant. Observations on species-specific habitat preferences showed An. stephensi to breed extensively in open wells and An. culicifacies in puddles/bed pools. Cx. tritaeniorhynchus was found breeding in almost all types of habitats surveyed. In retrospective, the study revealed information on mosquito species breeding in various habitats, which may be helpful for optimizing species sanitation. Keywords: breeding habitats, immature stages, vectors INTRODUCTION Rice is the staple food in India, and its cultivation has been traditionally carried out under different geographical and climatic conditions (Singh et al., 1989). Modern irrigation facilities and plant breeding technologies have resulted in increased acreage and cultivation of rice crops throughout the year leading to its increased production. Unfortunately, along with increased production it has also led to widespread outbreaks of mosquito borne diseases like malaria and Japanese encephalitis (JE; Lacey and Lacey, 1990). Corresponding author: Dr. K. John Ravindran, Integrated Disease Vector Control Project (NMRI), 332-A, Spartan School Road, Paneer Nagar, Mogappair, Chennai-600 037, Tamil Nadu, India. E-mail: johnravindran@gmail.com For their breeding, mosquitoes are known to exploit aquatic habitats present in riceland agro-ecosystem. Any change in this ecosystem, either naturally or man made, tend to alter the biotic community including mosquitoes, which may be advantageous or disadvantageous for mankind. Therefore, information on mosquito fauna and breeding ecology in a transitory environment is imperative for successful intervention. Ongoing urbanization in agricultural area in the outskirts of Chennai, Tamil Nadu, India, resulted in an ecologically transitional region, which received our attention. Information on breeding ecology of mosquitoes in this environment may be helpful to ensure preparedness for effective vector and disease control. 46 MATERIALS AND METHODS Chennai (longitude: 80° 15'E, latitude: 13° 05'N) isthe capital city of the state of Tamil Nadu, India, and is situated in the northeastern part of the state. Minimum mean and maximum mean temperatures recorded during 1990–2000 were 19.8° C and 38.5° C, respectively. Maximum rainfall is during Northeast monsoon (October–December). The study was undertaken in Chennai metropolitan area in western outskirts of the city during the period 1992–1999. Agricultural activities have been totally withdrawn in areas bordering the city due to ongoing urbanization and partially withdrawn in areas away from the outskirts of the city. The area represents an ecological transition zone. Studies on mosquito breeding in rice fields were carried out in agricultural fields located at Poonamallee and Thirukandalam areas, which are 10 km and 20 km away from the outskirts of Chennai. Observations were carried out from September 1992 to September 1993, covering one long-duration and two short-duration crops. Five fields were selected and regularly observed for mosquito breeding and population structure on a weekly basis during each crop. A quadrat with a dimension of 33 x 33 x 20 cm enclosing an area of 0.1m2 was used as a sampler (Chandrahas, 1990). Ten samples were taken/plot. Sampling was confined to previously earmarked sampling sites, which included four corner, four central and two random sites within 0.5 m from the bund. During each sampling occasion, height of rice plant, depth of standing water and peak noon water temperature were recorded with other information such as fertilizer used. In Irrigation wells, study period covered a premonsoon (August–October) and post-monsoon (November–February) period during 1998–1999. Observations were carried out in irrigation wells in riceland areas in Vanagaram area near Poonamallee, where irrigation wells suitable for the study were found. The conditions for selection of a well were absence of fishes, no floating vegetation and accessibility for sampling from all sides. Weekly observations were carried out in ten irrigation wells. A well net with a 20 cm diameter was used as a sampler. Immature samples were collected by pulling well nets through a distance of 1 m along the edges of the wall. Repeated sampling in the same area was avoided. Depth of water table and peak noon water temperature was recorded during each visit. Chi square analysis for Ravindran and Williams test of independence was performed to establish preferences towards breeding at various well depths. Habitat specific preferences were studied in few selected habitats namely open wells, bed pools, puddles and hoof-prints present in outskirts of periurban localities with or without ongoing agricultural activity. Any water stagnation with a surface area of less than 1 m2 was classified as a puddle and those with more than 1 m2 but less than 50 m2 as bed pool. Random surveys were undertaken and immatures were collected using well nets (wells), ladles (bed pools and puddles) and spoons (hoof-prints). Interspecific association and index of association in relation to breeding habitats of various mosquito vector species were calculated using the method of Cole (1949) and Whittaker and Fairbanks (1958). During sampling, immatures collected were counted instar-wise. Fourth instar larvae and pupae were brought to laboratory and reared until emergence for identification. RESULTS AND DISCUSSION A total of 11 Anopheles, 10 Culex and four Aedes species were collected from various breeding habitats in the study site (Table I). Among important disease vectors, An. stephensi and An. culicifacies, the urban and rural malaria vectors, respectively, Cx. tritaeniorhynchus, Cx. vishnui and Cx. pseudovishnui, the potential vectors of JE in peninsular Indian region and Ae. aegypti, vector of dengue were present. In rice fields, no malaria or filaria vectors were observed breeding. Similar results were obtained at Madurai (John Victor and Reuben, 1999) and Pondicherry (Chandrahas and Rajagopalan, 1979). Among vectors of JE, Cx. tritaeniorhynchus was predominant being encountered during all the 25 weeks of observation carried out over a period of three rice growing seasons, whereas Cx. vishnui and Cx. pseudovishnui were observed breeding during seven and one occasions only. Breeding pattern of Cx. tritaeniorhynchus differed from observations of Reuben (1971) and Rajendran and Reuben (1991) who reported intense breeding at the latter stages of plant growth. In Tamil Nadu, JE virus isolations have also been reported in An. subpictus, Cx. infula, Cx. whitmorei and Cx. fuscocephala, all of which breed in ricefields (Philip Samuel et al., 2000). Except Cx. infula and Cx. whitmorei, all other species were found breeding in rice fields. The density of immatures during the three cultivation 47 Mosquito breeding in riceland agro-ecosystem near Chennai seasons is given in Fig. 1 and 2. Anopheles and Culex density was high during first week after transplantation. Thereafter, intensity in breeding decreased. In Culex species, another peak in the density of immatures was observed during fourth to sixth week after transplantation. Standing water maintained for a period of two weeks after transplantation provided opportunity for intense exploitation of the habitat for breeding. Reduction in the density of immatures after this period, is due to alterations in daily irrigation practices that are controlled by various factors like soil texture, acreage and the number of crops raised in a year, erratic power supply and rainfall. Fields are usually irrigated immediately after cessation of standing water to manage adequate water supply. Such enforced intermittent irrigation due to lack of sufficient irrigation water has also been reported by Rajagopalan et al. (1990) and Russell et al. (1942) in areas in the adjacent erstwhile South Arcot district. In addition, rice cultivation practices such as deweeding, application of fertilizer and pesticides also reduce the density of immatures. In the present study, no pesticides were applied though application of urea as a fertilizer was undertaken before and three weeks after transplantation. 5000 4500 4000 3500 3000 2 Density/m 2500 2000 1500 1000 500 0 0 I II III IV VI V VII VIII IX X Weeks Transplantation Aug/Sep Jan/Feb Jun/Jul Fig. 1. Anopheles immature density in different rice growing seasons. 5000 4500 4000 3500 3000 2 Density/m 2500 2000 1500 1000 500 0 0 I II III IV VI V VII VIII Weeks Transplanta tion Aug/Sep Jan/Feb Jun/Jul Fig. 2. Culex immature density in different rice growing seasons. IX X 48 Ravindran and Williams Table I. Mosquito species observed breeding in different habitats in the study area Species Rice fields Irrigation wells Draw wells Puddles Hoofprints Bed pools Anophelines An. subpictus An. vagus An. peditaeniatus An. nigerrimus An. barbirostris An. stephensi An. aconitus An. splendidus An. annularis An. pallidus An. culicifacies + + + + - + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + Culicines Cx. tritaeniorhynchus Cx. vishnui Cx. pseudovishnui Cx. bitaeniorhynchus Cx. gelidus Cx. fuscocephala Cx. (Lutzia) fuscanus Cx. (Lophoceraomyia) sp. Cx. vegans Cx. quinquefasciatus Ae. vittatus Ae. scathophagoides Ae. aegypti Ae. vexans + + + + + + - + + + + + + + - + + + + + + - + + + + + + + + + + + + - + + + - Irrigations wells supported breeding of 15 species of mosquitoes. An. barbirostris, An. subpictus, An. vagus, Cx. quinquefasciatus and Cx. tritaeniorhynchus were potential breeders and were encountered breeding for more than 20 weeks. An. aconitus, An. splendidus, Cx. vishnui, Ae. vittatus, were very poor breeders and were obtained in less than five weeks during the course of observation. Among malaria vectors, An. stephensi was predominant being observed in 18 of the total 30 weeks of observation when compared to An. culicifacies, which was observed breeding only in two weeks and during postmonsoon season. Rapid urbanization in rice cultivation area adjoining extended areas may be the reason for prolific breeding of An. stephensi in these irrigation wells. Seasonal breeding preference in irrigation wells was noticed (Table II). An. aconitus and An. splendidus were found breeding in pre- monsoon season and An. culicifacies, An. nigerimus, Cx. vishnui, Cx. (Lophoceraomyia) species and Ae. vittatus were found breeding exclusively in postmonsoon season. All other mosquitoes were found breeding in both seasons. An. stephensi, An. subpictus, An. splendidus, An. Aconitus, Cx. quinquefasciatus and Cx. gelidus showed preference to breed at a well-depth greater than 15 feet, whereas An. barbirostris, An. vagus, An. nigerrimus, Cx. tritaeniorhynchus and Cx. (Lophoceraomyia) sp. at shallow levels with depth of less than five feet. Chi square analysis performed on species obtained at all well depths indicated preference of An. stephensi, An. barbirostris, An. vagus, Cx. quinquefasciatus and Cx. gelidus to breed at different well depths (Table III). Irrigation wells may act as foci for mosquito breeding during non-monsoon seasons and non-rice cultivation periods. 49 Mosquito breeding in riceland agro-ecosystem near Chennai Table II. Details of the density of immatures and larval emergence in irrigation wells Particulars Pre-monsoon Immature density/dip-Anopheles Immature density/dip-Culex Well Depth (Min.-Max.) Anophelines-No. emerged An. stephensi An. barbirostris An. vagus An. subpictus An. aconitus An. splendidus An. culicifacies An. nigerrimus Culicines-No. emerged Cx. quinquefasciatus Cx. tritaeniorhynchus Cx. gelidus Cx. fuscanus Cx. vishnui Cx. (Lophoceraomyia) sp Ae. vittatus 10.216.6 5.843.3 1417.3 918 210 (22.9) 431 (46.9) 34 (3.7) 238 (25.9) 2 (0.2) 3 (0.3) 0 (0)2 0 (0) 2544 1584 (62.3) 531 (20.9) 253 (9.9) 176 (6.9) 0 (0) 0 (0) 0 (0) Monsoon / Post-monsoon 12.722.2 5.323.9 2.623.2 1775 19 (1.1) 1094 (61.6) 322 (18.1) 285 (16.1) 0 (0) 0 (0) (0.1) 53 (3) 1282 437 (34.1) 145 (50.3) 15 (1.2) 34 (2.7) 41 (3.2) 106(8.3) 4 (0.3) (Figures in parenthesis denote percent) Table III. Mosquito breeding at different well depths Particulars No. of wells with mosquito breeding Anophelines An. stephensi An. barbirostris An. vagus An. subpictus An. aconitus An. splendidus An. culicifacies An. nigerrimus Culicines Cx. quinquefasciatus Cx. tritaeniorhynchus Cx. gelidus Cx. fuscanus Cx. vishnui Cx. (Lophoceraomyia) sp Ae. vittatus Well depth (ft) X2 <5 515 > 15 80 62 14 9 (11.3) 60 (86.3) 23 (28.8) 19 (23.8) 0 (0) 0 (0) 1 (1.3) 12 (15) 18 (29) 41 (66.1) 7 (11.3) 9 (14.5) 0 (0) 0 (0) 1 (1.6) 1 (1.6) 5 (35.7) 9 (64.3) 1 (7.1) 4 (28.6) 1 (7.1) 1 (7.1) 0 (0) 0 (0) 8.95* 9.04* 8.25* 2.44 NP NP NP NP 27 (33.8) 30 (37.5) 2 (2.5) 8 (10) 0 (0) 5 (6.3) 0 (0) 31 (50) 20 (32.3) 8 (12.9) 10 16.1) 1 (1.6) 0 (0) 4 (6.5) 9 (64.3) 4 (28.6) 2 (14.3) 1 (7.1) 0 (0) 0 (0) 0 (0) 6.62* 0.67 6.27* 1.59 NP NP NP (Figures in parenthesis denote percent; NP - not performed, * p > 0.05) - 50 Ravindran and Williams Table IV. Details of species specific habitat positivity and survey particulars Particulars Domestic wells Bed pools Puddles Hoof-prints No. surveyed No. positive for Anopheles breeding No. positive for Culex breeding Habitat positivity for breeding (%): An. subpictus An. vagus An. nigerrimus An. barbirostris An. stephensi An. aconitus An. annularis An. pallidus An. culicifacies Cx. tritaeniorhynchus Cx. vishnui Cx. bitaeniorhynchus Cx. gelidus Cx. fuscocephala Cx. (Lutzia) fuscanus Cx. (Lophoceraomyia) sp. Cx. vagans Cx. quinquefasciatus Ae. scathophagoides Ae. aegypti Ae. vexans 225 175 275 217 87 (59) 93 (56) 136 (74) 62 (35) 49 (27) 51 (33) 79 (45) 88 (65) 1.7 8.5 3.4 11.9 88.1 1.7 1.7 0 5.1 77.8 0 0 3.7 0 3.7 18..5 0 77.8 0 3.7 0 82.1 32.1 3.6 16.1 1.8 0 17.9 1.8 17.9 15.1 0 3 0 0 0 0 0 15.1 0 0 0 55.4 32.4 10.8 21.6 1.4 40 65.7 5.7 3 5.7 0 0 2.9 8.6 13.9 7.7 0 0 0 4.6 0 3.1 13.9 0 0 0 4.1 5.4 13.5 26.7 6.7 0 4.4 2.2 6.7 0 0 26.7 4.4 0 4.4 (Figures in parenthesis denote the number of habitats where adult mosquitoes emerged) Various mosquitoes sp. found breeding in puddles, bed pools, hoof-prints and domestic wells are shown in Table I. Puddles with rainwater were found to harbour the maximum number of species (16 nos.). Except for Ae. scathophagoides, all mosquitoes breeding in irrigation wells were also observed breeding in these habitats. An. stephensi was found to breed profusely in domestic wells and An. culicifacies in bed pools and puddles (Table IV). Breeding of An. stephensi in bed pools, hoof-prints and puddles is rather unusual but similar observations have been reported (Yadav et al., 1989). Breeding of An. stephensi in hoof-prints with rainwater collections in Chennai outskirts has been reported (Vasanthi, 1996). An. stephensi was predominant breeder in domestic wells, An. subpictus in bed pools and puddles and An. vagus in hoof-prints. Cx. tritaeniorhynchus, among JE vectors was most commonly encountered in all breeding habitats, whereas Cx. pseudovishnui was not encountered in any of the breeding habitats. Cx. vishnui was collected from puddles and hoof-prints only. Reuben (1971) reported breeding of Cx. tritaeniorhynchus and Cx. vishnui in wells, ponds, ditches, irrigation channels, borrow pits and from rice fields (fallow or planted) in North Arcot district of Tamil Nadu. Interspecific association (Table V) with regard to use of habitats for breeding of vectors (An. culicifacies, An. stephensi, Cx. tritaeniorhynchus and Cx. quinquefasciatus) existed between An. stephensi and An. culicifacies, and Cx. quinquefasciatus and Cx. tritaeniorhynchus in bed pools, indicating bed pools to be a very favourable source where all vector species of various diseases co-existed. In canal irrigated area 51 Mosquito breeding in riceland agro-ecosystem near Chennai Table V. Interspecific associations among important vector mosquitoes in study area Habitat Species Interspecific association (CAB) Index of association (I) Domestic well An. stephensi An. culicifacies An. stephensi - Cx. quinquefasciatus An. stephensi - Cx. tritaeniorhynchus An. culicifacies - Cx. quinquefasciatus An. culicifacies - Cx. tritaeniorhynchus Cx. tritaeniorhynchus - Cx. quinquefasciatus - 0.008 ± 0.314 -0.060 ± 0.146 -0.093 ± 0.167 0.036 ± 0.377 0.75 ± 1.445 0.012 ± 0.071 -0.970 -0.860 Bed Pools An. stephensi An. culicifacies An. stephensi - Cx. quinquefasciatus An. stephensi - Cx. tritaeniorhynchus An. culicifacies - Cx. quinquefasciatus An. culicifacies - Cx. tritaeniorhynchus Cx. tritaeniorhynchus - Cx. quinquefasciatus 1.000 ± 0.393 -0.833 ± 3.690 -0.970 ± 1.259 0.035 ± 0.079 0.737 ± 0.372 0.461 ± 1.526 -0.954 -0.848 Puddles An. stephensi An. culicifacies An. stephensi - Cx. quinquefasciatus An. stephensi - Cx. tritaeniorhynchus An. culicifacies - Cx. quinquefasciatus An. culicifacies - Cx. tritaeniorhynchus Cx. tritaeniorhynchus - Cx. quinquefasciatus -0.909 ± 2.795 -0.923 ± 2.519 -1.000 ± 0.786 -0.462 ± 0.64 -0.420 ± 0.370 0.180 ± 0.216 -1.000 -0.505 Hoof-Prints An. stephensi An. culicifacies An. stephensi - Cx. quinquefasciatus An. stephensi - Cx. tritaeniorhynchus An. culicifacies - Cx. quinquefasciatus An. culicifacies - Cx. tritaeniorhynchus Cx. tritaeniorhynchus - Cx. quinquefasciatus 0.600 ± 3.561 0.818 ± 1.98 0.111 ± 0.616 0.75 ± 1.591 0.938 ± 0.507 0.205 ± 0.277 -1.000 -0.762 in Gujarat, An. culicifacies was positively associated with An. stephensi only in paddy fields (Bhatt et al., 1990). Negative association was noticed in puddles, which was also similar to present findings. Likewise, in domestic wells a positive association existed between An. culicifacies, Cx. quinquefasciatus and Cx. tritaeniorhynchus. In puddles and hoof-prints, the coexistence of vectors was relatively low. Planned and systematic extension of Chennai city is underway resulting in rapid environmental changes. Both An. stephensi and An. culicifacies have been observed in extended regions of the city. An. stephensi, responsible for intense malaria transmission in Chennai dominated indicating species replacement on account of environmental change. Cx. tritaeniorhynchus, among potential vectors of JE was ubiquitous and was found to breed in all habitats surveyed. Intensive exploitation of rice fields throughout the rice growing season for breeding by this species is a cause of concern. However, natural enforced intermittent irrigation due to acute water shortage and withdrawal of rice cultivation may relatively contribute towards decreased risk in outbreak of JE in this area. Investigations on mosquito breeding in habitats in other extended areas may provide information on other species that may be prevalent in areas adjoining the city. Due to the prevalence of disease factor and high labour movement and activity, careful monitoring is indispensable for prevention of disease outbreaks. Bioenvironmental control, found to be successful in many areas, can be planned and adopted in this transition region before complete urbanization to achieve effective species sanitation. 52 ACKNOWLEDGEMENTS The authors wish to express their sincere thanks to Dr. V. P. Sharma and Dr. Sarala K. Subbarao, former Directors of National Institute of Malaria Research (NMRI), Prof. Dr. A. P. Dash, Director, NMRI and Shri Alex Eapen, Officer-in-charge, Integrated Disease Vector Control Project (IDVC; NIMR), Field Station, Chennai, for encouragement and facilities provided. The authors also thank Dr. R. C. Dhiman, Deputy Director, NMRI, for his critical review of the manuscript. The technical assistance rendered by staff of IDVC (NIMR), Field Station, Chennai, is also gratefully acknowledged. REFERENCES Bhatt RN, Sharma, RC and Kohli VK. 1990. Interspecific associations among anophelines in different breeding habitats of Kheda district Gujarat: Part ICanal Irrigated area. Indian J Malariol 27:167-172. Chandrahas RK. 1990. Host parasite interaction in ricefields and feasibility of mosquito control in urban areas. Ph. D. thesis, University of Berhampur, Orissa. Chandrahas RK and Rajagopalan P K. 1979. Mosquito breeding and the natural parasitism of larvae by a fungus, Coelomomyces and a mermithid nematode, Romanomermis, in paddy fields in Pondicherry. Indian J Med Res 69:63-70. Cole LC. 1949. Measurements of interspecific association. Ecology, 30:411-424. Dev V. 1994. Breeding habitats of anopheline mosquitoes in Assam. Indian J Malariol 31: 31-34. John Victor T and Reuben R. 1999. Population dynamics of mosquito immatures and the succession in abundance of aquatic insects in rice fields in Madurai, South India. Indian J Malariol 36:19-32. Ravindran and Williams Lacey LA and Lacey CM. 1990. The medical importance of rice land mosquitoes and their control using alternatives to chemical insecticides. J Amer Mosq Contr Assoc 6:1-93. Philip Samuel P, Hiriyan J and Gajana A. 2000. Japanese encephalitis virus infection in mosquitoes and its epidemiological implications. ICMR Bulletin 30:No.4. Rajagopalan P K, Das PK, Panicker KN, Reuben R, Raghunatha Rao D, Self LS and Lines JD. 1990. Environment and water management for mosquito control. In: Appropriate Technology in Vector Control. C F Curtis (Ed.) CRC Press, USA. pp 121-138. Rajendran R and Reuben R. 1991. Evaluation of the water fern Azolla microphylla for mosquito population management in the rice-land agro-ecosystem of south India Med & Vet Entomol 5:229-310. Reuben R. 1971. Studies on the mosquitoes of North Arcot District, Madras State, India. Part 5: Breeding places of the Culex vishnui group of species. J Med Ent 8:363-366. Russell P F, Knipe FW and Rao HR. 1942. On the intermittent irrigation of rice fields to control malaria in South India. J Mal Inst India 4:321-340. Singh Neeru, Singh OP and Soan V. 1989. Mosquito breeding in rice fields and its role in malaria transmission in Mandla district, M. P. Indian J Malariol 26:191-198. Vasanthi V. 1996. Field and laboratory studies on selected ecological and behavioural aspects of variants of Anopheles stephensi Liston from South India. Ph. D. thesis, University of Madras, Chennai. Whittaker R H and Fairbanks CW. 1958. A study of plankton copepod communities on the Columbian basin, Southeastern Washington. Ecology 39:46-65. Yadav R S, Sharma RC, Bhatt RM and Sharma VP. 1989. Studies on the Anopheline fauna of Kheda district and species-specific breeding habitats. Indian J Malariol 26:6574. Journal of Parasitic Diseases: June 2006, Vol. 30, No. 1, 53–57 J PD Random amplified polymorphic DNA of Trichomonas vaginalis isolates from Tarbiz, Iran 1 1 2 2 2 R. Jamali, B. Zareikar , A. Kazemi , M. Asgharzadeh , S. Yousefee , R. Estakhri , S. Montazer and A. 1 Ghazanchaei 1 2 Tarbiz University of Medical Science, Faculty of Medicine, Parasitology Department. Tarbiz University of Medical Sciences, Faculty of Medicine. ABSTRACT. Trichomonas vaginalis, the causative agent of human trichomoniasis, is the most common nonviral sexually transmitted disease. The infection may be asymptomatic or may cause severe vaginitis and cervicitis in women. Despite its high prevalence, little is known about its genetic variability and factors leading to asymptomatic infections. The random amplified polymorphic DNA (RAPD) technique is a simple method to detect DNA polymorphism. RAPD was performed by using four different random primers (OPD1, OPD2, OPD3 and OPD5) for the typing of 120 isolates of T. vaginalis from Tarbiz. Phylogenetic analysis was performed using SPSS program, and dendrogram with two distinct clusters was constructed. The asymptomatic isolates tended to form a cluster, separate from symptomatic isolates. Further studies for better understanding the relationship are suggested. Keywords: polymorphism, random amplified polymorphic DNA, Trichomonas vaginalis INTRODUCTION Trichomonas vaginalis is a prevalent vaginal pathogen, affecting 180 million persons worldwide annually (Wang 2000). Evidence from published studies exists that T. vaginalis is independently associated with a variety of adverse health consequences in both women and men, including increased human immunodeficiency virus (HIV) transmission, infertility, cervical intraepithelial neoplasia (CIN) development in women, and nongonoccocal urethritis and chronic prostatitis in men (Jane and Edward, 2003; Soper, 2004). Detection of T. vaginalis has traditionally relied on wet-mount microscopy or culture. These methods are highly specific but lack sensitivity. Nucleic acid Corresponding author: Dr. Rasoul Jamali, Parasitology Department, Faculty of Medicine, Tarbiz University of Medical Sciences, Daneshgah St, Tarbiz, Iran. Telefax : 0411-3364665, E-mail: rasooljamali@yahoo.com amplification assays are highly desirable alternatives to culturing, having both sensitivity and specificity for detecting T. vaginalis DNA (Schee et al., 1999). The random amplified polymorphic DNA (RAPD) technique represents as an efficient tool for the study of genetic polymorphism of DNA. It involves the amplification of random segments of genomic DNA by polymerase chain reaction (PCR) using short single primers of arbitrary sequences (Fraga et al., 2002). Different studies suggest that RAPD provides powerful markers to analyze the genetic diversity in T. vaginalis (Rojas et al., 2004; Kaul et al., 2004; Vanacova et al., 1997). The aim of this study was to genetically characterize by RAPD a collection of T. vaginalis isolates from patients with clinical signs and symptoms. MATERIALS AND METHODS A total of 2630 women visiting the health care centers of Tarbiz with and without symptoms of T. vaginalis 54 (vaginal discharge, itching, dysuria and dyspareunia) were selected and examined for the presence of T. vaginalis. From each individual two samples were collected from the posterior vaginal fornix by using two sterile cotton swabs. First swab was used for wet mount preparation and the second one was used to inoculate the Kupferberg medium (Quelab Laboratories, Canada). Culture tubes were incubated at 37° C up to 7 days, and examined microscopically on days 2, 3, 5 and 7 after inoculation. DNA extraction: Log phase T. vaginalis cultures were washed with phosphate buffered saline (pH 7.4) and the cell pellet was suspended in 400 µl TE (10 mM Tris, 1 mM EDTA) buffer (pH 8). To this suspension, 5 µl proteinase K (20 mg/ml) and 60 µl of 10% sodium dodecyl sulphate solutions were added, and incubated overnight at 55-65° C. Following incubation, 100 µl of NaCl and 80 µl of pre-warmed (at 65° C) CTAB/NaCl solution were added, vortexed well and incubated at 65° C for 10 min. DNA was cleaned by adding 700 µl of chloroform-isoamyl alcohol (24:1) solution and vortoxed for 20 s, and precipitated by 1ml of cold ethanol (70%) and centrifugation at 12,000 x g for 5 min (×2) at 10° C. Finally, after air-drying, the DNA pellet was dissolved in 50-100 µl TE buffer (pH 8). RAPD PCR: Four different 10 base pair primers were used for RAPD analysis (their sequence is shown in Table I; Snipes et al., 2000). The DNA amplification was performed at final volume of 25 µl containing: 2.5 µl of 10 x PCR reaction buffer (500 mM KCl and 200 mM Tris-HCl, pH 8.4), 1.25 µl MgCl2 (50 mM), 1 µl of each primer (Cinnagen, Iran), 0.5 µl of mixed dNTP (10 mM), 4 µl of template DNA, 15.35 µl of double distilled water and 0.4 µl of Taq DNA polymerase (5 unit/µl; Cinnagen, Iran). Negative controls for each of four primers used contained all components except template DNA. The amplification protocol consisted of an initial denaturation step at 94° C for 5 min followed by 40 cycle's repetitions of 1 min at 94° C, 1 min at 36° C and 2 min at 72° C. The final cycle “the extension step” was of 15 min at 72° C. The PCR products were analyzed by electrophoresis in 1.2% agarose gel in TBE buffer. The gels were then stained with ethidium bromide (0.5 µg/ml) and visualized under the UV transilluminator. RESULT Overall, 4.6% (120/2630) of specimens yielded a positive T. vaginalis culture. Seventy four isolates were obtained from symptomatic patients and 46 from Jamali et al. asymptomatic patients, axenically. DNA extracted from these isolates was subjected to RAPD analysis and amplified with 4 different random primers (Fig. 1a-d) show the RAPD patterns obtained with the primers used. All the primers provided distinct patterns. For each primer, the banding pattern was scored as presence (1) or absence (0) for each isolate and matrix table was constructed by SPSS 11.0 software. Dendrogram was built based on RAPD-PCR results for each primer, and for four primers, using Wards method and SPSS 11.0 program (Fig. 2). The isolates with similar banding pattern were assigned as a single type. OPD1 had the least typing ability as it gave 32 types for typing of 120 strains, whereas OPD5 had the highest typing ability that gave 58 types. OPD2 gave 56 and OPD3 gave 43 types. A total of 62 different types were obtained from 120 T. vaginalis specimens analyzed. There was one cluster consisting of four patients, seven with three patients, 41 with two patients and 13 with one patient. According to the tree, the isolates fell into two major groups (the classification results based on discriminant analysis are shown in Table II). The upper branch consisted of 65 isolates out of which 24 were from symptomatic patients and 41 isolates belonged to asymptomatic patients. The lower branch of tree consisted of 55 isolates, 50 of them from symptomatic patients and only five from asymptomatic ones. The 11 isolates that were from patients with the history of treatment failure, showed a scattered format in the tree (isolates 99, 83, 84, 70, 107, 27, 14, 7, 74, 9 and 36). Table I. The sequence of four primers used for RAPD analysis OPD OPD2 OPD3 OPD5 Size (mer) Squence (5' to 3') 10 10 10 10 ACCgCgAAgg ggACCCAACC gTCgCCgTCA TgAgCggACA Table II. Classification result based on discriminant analysisa Predicted group membership Ward method Original count 1 2 % 1 2 1 2 Total 62 3 3 52 65 55 95.4 5.5 4.6 94.5 100.0 100.0 a. 95% of original grouped cases correctly classified. 55 Random amplified polymorphic DNA analysis 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 Fig. 1-a: RAPD banding pattern of T. vaginalis isolates using OPD1 primer Lane 1: Size marker 100 bp DNA ladder Lane 2-23: Banding pattern of T. vaginalis isolates No.6 - 27 Lane 24: Negative control Lane 25: Size marker lambda DNA/EcoRI+HindIII 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 Fig. 1-b: RAPD banding pattern of T. vaginalis isolates using OPD2 primer Lane 1: Size marker 100 bp DNA ladder Lane 2-29: Banding pattern of T. vaginalis isolates No.75-102 Lane 30: Size marker lambda DNA/EcoRI+HindIII 1 2 3 4 5 6 7 8 9 10 11 12 Fig. 1-c: RAPD banding pattern of T. vaginalis isolates using OPD3 primer Lane 1: Size marker lambda DNA/EcoRI+HindIII Lane 2-13: Banding pattern of T. vaginalis isolates No.52-63 Lane 14: Size marker 100 bp DNA ladder 13 14 56 Jamali et al. 1 2 3 4 5 6 7 8 9 10 11 12 13 14 Fig. 1-d: RAPD banding pattern of T. vaginalis isolates using OPD5 primer Lane 1: Size marker lambda DNA/EcoRI+HindIII Lane 2-13: Banding pattern of T. vaginalis isolates No.52-63 Lane 14: Size marker 100 bp DNA ladder DISCUSSION Trichomoniasis is an important sexually transmitted disease which may manifest with a wide range of symptoms ranging from an asymptomatic presentation to severe sequel. It is `not clear yet as to why only a proportion of individuals infected with T. vaginalis become symptomatic, whereas the rest others remain asymptomatic. Strain variation and host factors may play a role in leading to symptomatic or asymptomatic infections (Kaul et al., 2004). Recent studies have shown the ability of DNA fingerprinting techniques in differentiating strains of various organisms (Tibayrenc, 1998). Vanacova et al. (1997) for the first time used RAPD technique for phylogenetic analysis of T. vaginalis and found it a useful method in epidemiological analysis. Their results suggested a concordance between the genetic markers with resistance to metronidazole and clinical findings, but they found no concordance with the presence of T. vaginalis virus (TVV) and the virulence of strains. Hample et al. (2001) assayed the relationship between 20 strains of T. vaginalis from eight countries using RAPD analysis, and they found that the phylogenic tree reflects the pattern of virulence, geographic origin or infection by TVV. Rojas et al. (2004) used RAPD technique in 40 isolates of T. vaginalis to find an association between genetic polymorphism of organism and its clinical characters. Their results emphasize that the severity of infection depends on the genetic type of T. vaginalis involved. In the present study, we used RAPD method for the genetic analysis of 120 clinical isolates of T. vaginalis, and we investigated the association of T. vaginalis genetic polymorphism and its clinical classification as symptomatic or asymptomatic. In conclusion, our results show that the isolates from asymptomatic patients tend to form a distinct cluster separate from symptomatic isolates, and that T. vaginalis isolates from patients with or without symptoms are genetically different. Further studies are necessary to better understand the relationship between genetic markers and the pathogenicity of the organism. REFERENCES Fraga J, Rojas L, Sariego I and Sarria CA. 2002. Optimization of random amplified polymorphic DNA technique for its use in genetic studies of Tricomonas vaginalis isolates. Infect Genet E 2: 73-75. Hample V, Vanacova S, Kulda J and Flegr J. 2001. Concordance between genetic relatedness and phenotypic similarities of Trichomonas vaginalis strains. BMC Evol Biol 48:1-11. Jane RS and Edward W. 2003. High rate of Trichomonas vaginalis among men attending a sexually transmitted Random amplified polymorphic DNA analysis Rescaled Distance cluster Combine C A S E 0 5 10 15 20 25 Label Num 89 57 disease clinic: implications for screening and urethritis management. J Infect Dis 188: 465-468. 90 91 39 68 32 115 99 101 100 94 Kaul P, Gupta I, Sehgal R and Malla N. 2004. Trichomonas vaginalis: random amplified polymorphic DNA analysis of isolates from symptomatic and asymptomatic women in India. Prasitol Int 53: 255-268. 98 95 96 93 80 83 87 102 Rojas L, Fraga J and Seriego I. 2004. Genetic variability between Trichomonas vaginalis isolates and correlation with clinical presentation. Infect Genet E 4:53-58. 88 81 82 84 111 113 55 119 106 45 70 103 Schee C, Belkum A, Zwijgers L, Brugge E, Oneill EL, Luijedjk A et al.1999. Improved diagnosis of Trichomonas vaginalis infection by PCR using vaginal swabs and urine specimen compared to diagnosis by wet mount microscopy, culture and fluorescent staining. J Cli Microbiol 37:4127-4130. 110 104 105 109 107 107 114 120 46 61 Snipes LJ, Gamard PM, Narcisi EM, Ben Beard C, Lehman T and Secor EW. 2000. Molecular epidemiology of metronidazole resistance in a population of Trichomonas vaginalis clinical isolates. 38: 3004-3009. 116 97 112 28 29 24 Soper D. 2004. Trichomoniasis:under control or undercontrolled. Am J Obstet Gynecol 190: 281-290. 44 60 27 1 21 52 12 14 Tibayrenc M. 1998. Beyond strain typing and molecular epidemiology: integrated genetic epidemiology of infectious disease. Parasitol Today 14: 323-329. 26 108 30 66 71 49 86 34 Vanacva S, Tachezy J and flegr J. 1997. Characterization of trichomonad species and strains by PCR fingerprinting. J Eukayot Microbiol 44: 545-552. 37 118 11 13 3 4 6 7 5 10 16 59 17 2 18 15 9 67 77 62 69 64 58 72 73 74 54 56 76 63 75 48 51 50 23 57 38 40 41 42 47 25 43 65 78 79 9 36 85 33 53 31 35 19 20 22 92 Fig. 2. Dendrogram for 120 isolates of Trichomonas vaginalis based on RAPD-PCR data. Wang J. 2000. Trichomoniasis. Prim Care Update Ob/Gyns 7:148-153. Journal of Parasitic Diseases: June 2006, Vol. 30, No. 1, 58–63 J PD Two new species of Trypanosoma from freshwater fish (Heteropneustes fossilis and Channa punctatus) from Bareilly, India 1 D. K. Gupta, N. Gupta and R. Gangwar Department of Zoology, Bareilly College, Bareilly. Department of Animal Science, M. J. P. Rohilkhand University, Bareilly. 1 ABSTRACT. Haematological examination of fresh water fish Channa punctatus (115 specimens) and Heteropneustes fossilis (163 specimens) revealed 10.6% and 14.6% infection, respectively, of dimorphic species of Trypansoma. Both in the former and latter hosts, the parasite concentration index was 112.4 and 110.5 Trypanosomes/100 RBCs, respectively. The flagellate from C. punctatus is characterized by its dimorphic nature [small (18-33 µm) and large (30-52 µm) forms], and was found to be distinct from the earlier reported species in total body length, flagellar length, kinetoplast dimensions and granulation pattern. The trypanosome species from H. fossilis was also observed to be dimorphic [small (23.5-30.5 µm) and large (33.1-43.8 µm) forms], and differences existed in cell breadth, nuclear size, flagellar length and cytomorphological features from the earlier recorded species. These two dimorphic species have been described herein as Trypanosoma saulii n. sp and Trypanosoma heteropneusti n. sp from C. punctatus and H. fossilis, respectively. Keywords: Channa, dimorphic, division, Heteropneustes, Trypanosoma INTRODUCTION The freshwater fish, Channa punctatus and Heteropneustes fossilis have served as favourable hosts of Trypanosoma: T. ophiocephali Pearse, 1933; T. striati Qadri, 1955; T. punctati Hasan and Qasim, 1962; T. elongatus Raychaudhuri and Misra, 1973; T. bareilliana Gupta et al., 1987 and T. rohilkhandae Gupta and Saraswat, 1991 have been reported from various species of Channa. The dimorphic species recorded are T. gachuii Misra et al., 1973 from C. gachua and T. aligaricus Gupta and Jairajpuri, 1982a from C. punctatus. On the other hand, T. saccobranchi Castellani and Willey, 1905; T. danilewskyi Qadri, 1962; T. singhii Gupta and Jairajpuri, 1981; T. Corresponding author: Prof. Neelima Gupta, Department of Animal Science, M.J.P. Rohilkhand University, Bareilly 243 006, U.P., India. E-mail: guptagrawal@rediffmail.com kargenensis Gupta and Gupta, 1994 and T. karelensis Gupta et al., 2001 have been recorded from H. fossilis; however, T. mukundi Raychaudhuri and Misra, 1973 is the only dimorphic species. Herein, two new species of Trypanosoma have been recorded from C. punctatus and H. fossilis. MATERIALS AND METHODS Live specimens of C. punctatus and H. fossilis, collected from fresh waters of Bareilly were transported to the laboratory and maintained in separate aquaria under optimum conditions of food and aeration. Blood was collected from their caudal vein and examined immediately (hanging drop preparation and micro-haematocrit) for the presence of parasites. When positive, smears were made, air dried, fixed in methanol and stained in Leishman + phosphate buffer (pH 6.6) in the ratio of 1:7 for 40 min. 59 Two new species of Trypanosoma After thorough washing, the films were air dried, mounted in DPX, sealed and observed and photographed at 1000 x magnification under LEICA DMLB photoautomat. Camera lucida drawings were made for statistical measurements of the parasites. Nuclear index (NI), flagellar index (FI) and kinetoplast index (KI) were calculated as follows: NI = PN/NA; FI = CL/FL; KI = PN/KN where PN = posterior end to nucleus ; NA = anterior end to nucleus ; CL = body length ; FL = flagellar length and KN = kinetoplast to nucleus. RESULTS Trypanosoma saulii n.sp. (Fig. 1 A-D; Table 1) Host - Channa punctatus Bloch Locality - Bans Mandi, Bareilly, India Percentage of infection/intensity - 10.6%/1-12.4 Trypanosomes/100 RBCs (n = 115) The live parasite in hanging drop preparation showed an active wriggling movement amongst the blood corpuscles. Out of the infected fish, 6.4% were moderately infected (parasitaemia 5-12.4 Trypanosomes/100 RBCs), the remaining had scant infection (parasitaemia 1-5 Trypanosomes/100 RBCs). In stained smears, two forms of the parasite could be differentiated : minuta and magna. Kinetoplast: Well developed structure and takes deep stain upon fixation. Shape may be conical or spherical. Division of the kinetoplast not observed. Flagellum and undulating membrane: Flagellum originates from kinetoplast, runs anteriorly bordering the undulating membrane and finally extends beyond body as a free flagellum. Flagellum well-developed in larger forms than smaller forms. Undulating membrane conspicuous and takes a light stain. It extends throughout length of cell body. In smaller forms, it has about 1-2 folds, whereas in larger forms it has 1-3 folds. Width of undulating membrane greater in larger forms. Indices FI KI NI = = = 1.92 1.72 1.62 DISCUSSION The genus Channa (=Ophiocephalus) has, from time to time, been reported as a suitable host for protozoan parasites (Haldar and Mukherjee, 1979). Various trypanosome species have been put on record and a comparison indicates that T. ophicephali, T. punctati and T. elongatus are monomorphic forms and, A B C 1. Morphology Shape: Trypanomastigote elongated, anterior and posterior ends tapering, the latter may assume a beaklike appearance. Configuration of parasite usually of letter 'C', sometimes elongated 'S' and in rare cases, almost a straight line. Posterior end extremity in both (small and large) cases, more or less rounded. Cytoplasm: Granular cytoplasm stains lightly. In the smaller forms, the granules more concentrated in the anterior region of the body commencing from just anterior tip of the nucleus to approximately half the distance from their commencement to the origin of the flagellum. In large forms, granules occur along both sides of nucleus. Nucleus: Deep red stained nucleus oblong or beanshaped in middle of cell body, occupying almost entire cell breadth. D E F G H J I K 10 µm Fig. 1. A-D T. saulii n. sp. (A, B: minuta froms; C, D: magna forms), E-K T. heteropneusti n. sp. (E, F: smaller forms; G, H: larger forms; I-K: diving forms) 60 Gupta et al. therefore, the present form is distinct from them. The present species also can not be compared to T. striati, T. channai and T. bareilliana which are polymorphic species. The dimorphic species recorded from C. punctatus are T. gachuii and T. aligaricus (Table I) and the present form warrants comparison with these species. A critical examination indicates that the trypanosome described herein is not comparable to both of these species in the total body length (CF) as the present form is a much smaller species, the larger form approximates the smaller form of the above mentioned species. Again, the length of the cell body of the parasites is also not comparable as also the flagellar length (FL). Moreover, the present form is also not comparable with T. gachuii in breadth of cell body (CB) although it may be comparable to T. aligaricus. Interestingly, although the parasite is much smaller than T. gachuii and T. aligaricus but the nuclear length (NL) is comparable to T. aligaricus indicating a relatively larger nuclear size in relation to its total length. The kinetoplast dimensions of T. gachuii have not been provided by the authors but the kinetoplast of the present form is again comparatively larger than in T. aligaricus. Apart from the above statistical differences, the present parasite also differs from the compared species in the granulation and vacuolation pattern. Although two other species have been described from the same ecological niche, the present form cannot be synchronized with them due to its dimorphic nature. infected fish, 4.2% were moderately infected (parasitaemia examined 5-10.5 trypanosomes/100 RBCs) and the remaining had scant infection (parasitaemia 1-5 trypanosomes/100 RBCs). In stained smears, the smaller and larger forms could be clearly identified showing dimorphism. MORPHOLOGY Shape: 'C' or 'S' shaped body somewhat elongated but usually takes twisted conformation upon fixation. Cytoplasm: Lightly stained granular cytoplasm; granules more prominent in posterior part of parasite. Vacuoles were absent. Nucleus: Deeply stained oval or bean-shaped nucleus situated in the middle of body. Nucleolus not observed. Kinetoplast: Oval-shaped kinetoplast posteriorly placed, takes a deep stain upon fixation. Flagellum and undulating membrane: Flagellum originates from kinetoplast. Free-flagellum moderately well developed. Light pink stained undulating membrane has 2-4 folds; 1-2 major folds may also be present. mi Ia sm mg The above discussion indicates that the present form is different from any species reported either from the same ecological niche or elsewhere. It is, therefore, proposed that based on the geographical location, morphometrics, cytological pecularities and host status, the species discovered from the blood of C. punctatus collected from Bareilly region, India, should be given a new species status and the name T. saulii n. sp is proposed. Trypanosoma heteropneusti n.sp. (Fig. 1 E-H; Table II) Host Locality Percentage of infection/intensity - Heteropneustes fossilis - Local fish market, Bareilly - 14.6%/1-10.5 Trypanosomes/100 RBCs (n = 163) The live parasites showed an active wriggling movement amongst the blood corpuscles. Out of the E Fig. 2. Photomicrographs of Trypanosoma. A. T. saulii n. sp. (miminuta form; mg - magna form), B-F. T. heteropneusti n.sp. B. dimorphic forms (sm-small form, la - large form), C. differentiating nucleus, D. longitudinal kinetosome division, E and F. migrating nuclei. 61 Two new species of Trypanosoma Indices FI KI NI = = = 1.82 1.72 1.16 DISCUSSION H. fossilis has been considered to be an ideal host for trypanosomes. A dimorphic form, T. mukundi was discovered by Raychaudhri and Misra (1973) from the fresh waters of Kolkata, India existing as “slender” and “stumpy” forms. Table II shows a comparison of dimorphic parasites recorded from the test fish. T. mukundi Raychaudhuri and Misra (1973) is a dimorphic species and the present form warrants a critical examination of the said species. The contour of the parasites differ, the present form being “slender” and “elongated”, whereas T. mukundi has been reported as “slender” and “stumpy” form. The present forms have therefore contrastingly been described as “small” (23-30.5 µm) and “large” (33.1-43.8 µm) forms. Although CL of both parasites are comparable but conspicuous difference in CB and nuclear size are evident. Moreover, the flagellar lengths of both the parasites also differ. The granulation pattern and the vacuolation of both the parasites cannot be treated at par. It is, therefore, proposed that the parasite discovered from the blood of H. fossilis collected from the fresh waters of Bareilly be designated as a new species and, accordingly, the name T. heteropneusti n.sp is proposed to accommodate the parasite with the specific characters as given in this account. DIVISION IN T. heteropneusti N.SP. Some forms undergoing division showed peculiar features in the blood of H. fossilis. However, only two fish out of 163 specimens exhibited multiplying forms. Division commenced in the nucleus showing well differentiated chromatin prior to division (Fig. 1I). The two centrally-placed, round to ovoid nuclei and the single, undivided kinetoplast was placed Table I: Dimorphic trypomastigotes from Channa (= Ophiocephalus; all measurements in µm) Host Parasite Author(s) Component parts/forms* O. gachua T. gachuii Misra et al., 1973 C. punctatus T. aligaricus Gupta and Jairajpuri, 1982 C. punctatus T. saulii : n.sp (N=50) Present study Short Slender Small Large Minuta Magna CF 36.4 53.9 34.37 (34.25-35.5) 54.68 (53.25-55.3) 24.0 (18.0-33.0) 35.5 (30.2-52.0) CL 24.9 42.1 23.50 (23.01-25.0) 46.18 (44.95-46.8) 18.5 (12.2-24.4) 28.2 (26.3-36.4) CB 2.3 3.9 1.65 (1.5-1.8) 2.45 (2.1-2.6) 1.9 (1.0-2.5) 2.6 (2.0-3.6) FL 11.1 10.9 10.87 (10.25-13.5) 8.50 (7.82-9.4) 6.6 (4.0-8.2) 8.8 (8.0-12.0) NL 2.4 2.7 3.00 (2.5-3.5) 3.73 (3.6-3.8) 3.2 (1.0-6.0) 3.7 (1.0-6.0) NB 2.2 2.7 1.12 (1.0-1.2) 2.02 (1.8-2.2) 1.6 (1.0-3.0) 2.0 (1.0-4.0) KB - - 0.45 (0.4-0.5) 0.25 (0.21-0.3) 1.2 (0.5-2.0) 1.25 (0.5-2.0) KL - - 10.5 (0.5-1.6) 0.51 (0.4-0.6) 1.6 (1.0-2.0) 1.9 (0.5-2.0) * CB= Breadth of cell body; CF = Total length; CL = Length of cell body; FL = Flagellar length; KB = Kinetoplast breadth; KL = Kinetoplast length, NB = Nuclear breadth; NL = Nuclear length 62 Gupta et al. Table II: Dimorphic trypomastigotes from Heteropneustes fossilis (all measurements in µm) Parasite Author(s) Locality Component Parts/forms T. mukundi Raychaudhuri and Misra 1973, India T. heteropneusti n.sp Present study India (n = 50) Slender Stumpy Small Large CF 32.5-43.5 22.0-28.0 27.9 (23.5-30.5) 37.83 (33.1-43.8) CL 24.0-28.5 18.0-23.5 16.7 (14.4-19.0) 22.6 (20.1-24.2) CB 2.0-2.5 1.9-2.5 1.68 (1.45-1.95) 1.40 1.3-1.5) FL 9.0-15.5 4.0-6.5 8.48 (6.95-11.4) 16.05 (12.05-18.6) NL 2.25-3.75 2.25-3.75 2.64 (1.41-3.35) 2.63 (1.51-3.30) NB 1.75-2.25 1.75-2.25 1.14 (1.1-1.2) 1.23 (0.95-1.25) KB 0.75-7.5 0.75 0.93 (0.90-0.95) 0.90 (0.8-0.95) KL - - 1.18 (1.07-1.35) 1.16 (0.8-1.35) * Abbreviations same as in Table I posteriorly with a single, long free-flagellum and a normal undulating membrane (Fig. 1J). The kinetoplast usually divided subsequent to nuclear division but in some instances, it was the first to divide. Interestingly, both longitudinal as well as transverse division occurred during the division of kinetoplast although the former was more frequent (Fig. 2D). The two nuclei now migrated, one in either arm (Fig. 2E) and the parasite stretched between the two kinetoplasts prior to cytokinesis (Fig. 2F). One abnormal trypanosome with three kinetoplasts (Fig. 1K), dividing prior to nuclear division, could also be observed which does not fit into the sequence of divisional events of trypanosome multiplication and is apparently an aberrant form. Division in trypanosomes has been reported rarely. Laveran and Mesnil (1907) reported absence of division in T. remarki but occasionally observed forms with two nuclei in the parva variety. T. danilewskyi divided at 20° C satisfactorily in gold fish but the process slowed down by increasing (30° C) or decreasing (10° C) the temperature (Lom, 1973; Woo et al., 1983; Islam and Woo 1992). Qadri (1955), Daly and Degiusti (1971), Khan (1972), Grogl et al. (1980) and Mukerjee and Haldar (1982) reported the absence of divisional stages in the fish blood. Longitudinal binary fission in culture form of T. striati (Qadri, 1955); binucleated trypomastigotes in T. batrachi Qadri (1962), T. maguri and T. vittati (Tandon and Joshi, 1973), T. mrigali (Joshi, 1976) and T. attii (Gupta and Jairajpuri, 1982b) have also been observed. Two small trypanosomes possessing two nuclei and two kinetoplasts (Baker, 1960), and two opposite nuclei and a central kinetoplast (Raychaudhuri and Misra, 1973) have also been put on record. Wenyon (1926) and Lewis and Ball (1981) reported kinetoplast division prior to nuclear, whereas Burreson (1982) recorded a reverse condition in Trypanoplasma bullocki. In T. heteropneusti n.sp, both instances have Two new species of Trypanosoma been recorded but prior nuclear division dominates prior kinetoplast division. The nuclear division in our specimens indicate only the transverse division. The cytokinesis and flagellar division were not observed. ACKNOWLEDGEMENT Thanks are expressed to the University Grant Commission, New Delhi, India, for providing financial grant in the form of a research project. REFERENCES Baker JR. 1960. Trypanosomes and dactylosomes from the blood of freshwater fish in East Africa. Parasitology. 50 : 515 - 526. Burreson EM. 1982. The life cycle of Trypanoplasma bullocki (Zoomastigophorea: Kinetoplastida). Journal of Protozoology. 29 : 72 - 77. Castellani A and Willey A. 1905. Observations on haematozoa in Ceylon. Quarternary Journal of Microscopic Science. 49 : 383 - 402. Daly JJ and Degiusti DL. 1971. Trypanosoma catostomi n. sp. from the white sucker Catostomus C. commersoni. Journal of Protozoology.18 : 414 - 417. Grogl M, Marinerlle CJ, Suarez MF, Sanchez NH and Guhl F. 1980. Trypanosoma magdalenae n. sp. (Protozoa: Kinetoplastida) from a fresh water teleost, Petenia kraussii in Columbia. Journal of Parasitology. 66 : 1022 - 1026. Gupta DK and Gupta N. 1994. A comparative study of culture media for Trypanosoma kargenensis n. sp. from Heteropneustes fossilis and its effect on the biochemical composition of fish. In: Proc. Third Asian Fish. Forum. Chou et al. (Eds) A. F. S. Philippines. pp320-323. Gupta DK, Gupta N and Yadav P. 1987. New polymorphic trypanosomes from two freshwater fishes. Himalayan Journal of Environment and Zoology. 1 : 93 - 97. Gupta N, Gupta DK and Saraswat H. 2001. Hypoglycemia in Heteropneustes fossilis parasitized by two new species of parasites Trypanosoma karelensis n.sp and Myxosoma fossilii. n.sp. In: Proceedings of the National Symposium on Fish Health, Management and Sustainable Aquaculture.pp179-186. Gupta N and Jairajpuri DS. 1981. Trypanosoma singhii n. sp. from a fresh water teleost, Heteropneustes fossilis. Indian Journal of Parasitology. 5 : 251 - 253. Gupta N and Jairajpuri DS. 1982a. Trypanosoma aligaricus n. sp. from the freshwater murrel, Ophiocephalus punctatus Bloch. Archiv fur Protistenkunde. 125 : 109 - 114. Gupta N and Jairajpuri DS. 1982b. A new polymorphic trypanosome from an Indian freshwater fish Wallago attu (Bl. and Schn.) Rivista di Parassitologia. 43: 287-292. Gupta N and Saraswat H. 1991. Trypanosoma rohilkhandae n. sp. from fresh water teleost fish Channa (= Ophiocephalus) punctatus. Himalayan Journal of Environment and Zoology. 5 : 29 -33. 63 Haldar DP and Mukherjee M. 1979. An annolated list of protozoan parasites from Ophicephalus punctatus Bloch, a common food - fish. Archiv fur Protistenkunde. 121 : 392 400. Hasan R and Qasim SZ. 1962. Trypanosoma punctati n. sp. from the fish Ophiocephalus punctatus Bloch a common fresh water murrel of India. Zeitschrift fur Parasitenkunde. 22 : 118 - 122. Islam AKMN and Woo PTK. 1992. Effects of temperature on the in vivo and in vitro multiplication of Trypanosoma danilewskyi, Laveran and Mesnil. Folia Parasitologica. 39 : 1 - 12. Joshi BD. 1976. Two new species of trypanosomes from fresh water teleosts. Indian Journal of Zootomy. 17 : 5 - 10. Khan RA. 1972. On a Trypanosoma from the Atlantic cod, Gadus morhua. Canadian Journal of Zoology. 50 : 1051 1054. Laveran A and Mesnil F. 1907. Trypanosomes and Trypanosomiasis. Ist Ed. Bailliere, Tindall and Cox (Translated into English by D. Nabarro) London. Lewis JW and Ball SJ. 1981. Observation on the division of epimastigotes of Trypanosoma cobitis. Archiv fur Protistenkunde. 124 : 337 - 344. Lom J. 1973. Experimental infection of gold fish with blood flagellates. In : Progress in Protozoology, Proceedings of the International Congress in Protozoology. pp 255. Misra KK, Chandra AK and Choudhury A. 1973. Trypanosoma gachuii n. sp. from a fresh water teleost fish Ophicephalus gachua Ham. Archiv fur Protistenkunde. 115: 18 - 21. Mukherjee M and Haldar DP. 1982. Host susceptibility experiments on a new fish trypanosome in India. Angewandte Parasitology. 23 : 69 - 73. Pearse AS. 1933. Parasites of Siamese fishes and crustaceans. Journal of the Siamese Siena Society of Natural History, Supplement. 9 : 179 - 191. Qadri SS. 1955. The morphology of Trypanosoma striati n. sp. from an Indian freshwater fish. Parasitology. 45 : 79 - 85. Qadri SS. 1962. An experimental study of the life cycle of Trypanosoma danilewskyi in the leech, Hemiclepsis marginata. Journal of Protozoology. 9 : 254 - 258. Raychaudhuri S and Misra KK. 1973. Two new fish trypanosomes from India. Archiv fur Protistenkunde. 115 : 10 - 17. Tandon RS and Joshi BD. 1973. Studies on the physiopathology of blood of freshwater fishes infected with two new forms of trypanosomes. Zeitchrift fur Wissen schaftliche Zoologie. 185 : 207 - 221. Wenyon CM. 1926. Protozoology. Bailliere, Tindall and Cox. London. pp1563. Woo PTK, Wehnert DS and Rodgers D. 1983. The susceptibility of fishes to haemoflagellates at different ambient temperatures. Parasitology. 87 : 385 - 392. Journal of Parasitic Diseases: June 2006, Vol. 30, No. 1, 64–67 J PD Impact of anthelmintic therapy on live weight gain in gastrointestinal nematode-infected goats* A. K. Jayraw and Y. V. Raote Department of Parasitology, College of Veterinary and Animal Sciences, Parbhani. ABSTRACT. A trial was conducted to assess the effect of anthelmintic therapy on live weight gain in gastrointestinal (GI) nematode-infected goats using three anthelmintics, viz. tetramisole hydrochloride, morantel citrate and albendazole. Twenty four goats with heavy natural infection of Strongyle, Strongyloides papillosus and Trichuris spp. were selected and divided into four groups of six animals each. Group-I served as untreated control group, whereas Group-II, III and IV were treated with tetramisole hydrochloride @ 15 mg kg-1 body weight (b.w.), morantel citrate @ 5.94 mg kg-1 b.w. and albendazole @ 7.5 mg kg-1 b.w., respectively. Animals were weighed at weekly intervals and their live weight was recorded for a period of 28 days post-treatment (PT). On 28 day PT, overall live weight gain recorded was 9, 7.44 and 8.78% in animals of Group-II, III and IV, respectively, whereas only 3.73% weight gain was recorded in goats of untreated control group. The net profit obtained by the end of experimental study was Rs. 206, 138 and 174 in animals of GroupII, III and IV, respectively, as against a profit of Rs. 94 obtained in animals of untreated control group. Keywords: anthelmintic therapy, gastrointestinal nematode, goat, weight gain INTRODUCTION Owing to the growing demand for high quality animal proteins for human consumption, small ruminants occupy a special place as they are extremely efficient in converting the indigestible cellulose and hemicellulose to animal protein. India has a population of 120 million goats, which contribute greatly to rural economy (Abraham, 2001). Control of gastrointestinal (GI) nematodosis is one of the most serious challenges, as it poses a major constraint in the growth of highly profitable animals due to the chronic and insidious nature of parasitism (Sanyal, 1996). In *Part of M.V. Sc. thesis by first author, Marthwada Agricultural University, Prabhani. Corresponding author: Dr. A. K. Jayraw, Department of Parasitology, Nagpur Veterinary College, Nagpur-440 006, Maharashtra, India. the absence of a potent prophylactic agent, chemotherapy continues to play a vital role against GI nematodosis. Therefore, the present investigation was undertaken to evaluate the live weight gain response in goats following chemotherapy with commonly available anthelmintics, viz. tetramisole hydrochloride, morantel citrate and albendazole. MATERIALS AND METHODS The experimental study was conducted at the College of Veterinary and Animal Sciences, Parbhani during monsoon. A total of 49 female goats were maintained under a semi-intensive system and each animal was provided with 250 g concentrate daily. The helminthic burden of each animal was ascertained after counting eggs per gram (EPG) of faeces using Stoll's dilution method (Soulsby, 1982). Out of 49 goats, 1-2 year old naturally infected 24 animals were selected, showing Effect of antihelmenthic therapy on live weight gain the mean EPG counts of 3783 ± 1106.96; 404.16 ± 127.54 and 212.5 ± 55.01 for strongyle species, S. papillosus and Trichuris spp., respectively. The experimental animals were equally divided into four groups (Gr.), where, Gr.-I served as infected, untreated control group, Gr.-II animals were treated with tetramisole hydrochloride (Nilverm soluble powder, M/S ICI India Ltd., Kolkata) @ 15 mg kg-1 body weight (b.w.) and Gr.-III animals with morantel citrate (Banminth Tab., M/S Pfizer Ltd., Ahmedabad) @ 5.94 mg kg-1 b.w. The animals in Gr.-IV were drenched with albendazole (Albomar suspension, M/S Glaxo India Ltd., Mumbai) @ 7.5 mg kg-1 b.w. The experimental goats were denied access to water and feed for approximately 18 h, prior to recording their live weight (weekly) using a weigh-bridge. The net profit in terms of rupees (Rs.) was calculated taking into account the final difference in body weight of the experimental goats before and after chemotherapy, cost of medication including cost of drug (tetramisole hydrochloride - Rs. 10, morantel citrate - Rs. 29 and albendazole - Rs. 24) and labour cost (Rs. 60 per head for one day), except for the animals belonging to Gr.-I, which served as infected untreated control and were not treated with any anthelmintic. The average rate of chevon was considered as Rs. 120 kg-1 at Parbhani district of Maharashtra. The data obtained were analysed following standard statistical procedures (Snedecor and Cochran, 1994). RESULTS Animals were weighed at weekly intervals and their live weight recorded for a period of 28 days. The mean EPG counts recorded at 28 days post-treatment were found to be 345.84 ± 290.47, 4.165 ± 4.165 and 154.16 ± 37.50 for strongyle species, S. papillosus and Trichuris spp., respectively. A maximum of 9% (mean 1.97 kg) live weight gain was recorded in tetramisole hydrochloride treated animals (Gr.-II) followed by 8.78% enhancement (mean 1.82 kg) in experimental goats belonging to albendazole treated group (Gr.-IV). The morantel citrate treated group (Gr.-III) exhibited 7.44% weight gain (mean 1.56 kg). The lowest weight gain of 3.73% (mean 0.79 kg) was evident in animals of infected untreated group (Gr.-I). The mean daily weight gain recorded is presented in the descending order as 70 g day-1 in animals of Gr.-II, 65 g day-1 in Gr.IV, 55 g day-1 in Gr.-III and 28 g day-1 in animals of Gr.I. The net profit obtained at the end of experimental study was higher (Rs. 206) in tetramisole hydrochloride treated group, when compared with 65 albendazole (Rs.174) and morantel citrate (Rs. 138) treated groups. On the other hand, the net profit recorded in infected untreated control group was Rs. 94 in comparison with the treated groups (Table I). DISCUSSION GI nematode infection is recognized as a major constraint to small ruminant production worldwide. It is well known to cause huge economic losses due to mortality incidental to the severe parasitism and high morbidity, reduction and/or spoilage of wool, meat and milk production in chronic cases (Chowdhury, 1994). The continued presence of GI nematodes is responsible for anorexia, reduced feed intake, alterations in protein metabolism, low levels of minerals, depressed activity of some intestinal enzymes and diarrhoea (Soulsby, 1982). The parasitism is also associated with loss of blood and plasma proteins into the GI tract (Jayraw and Raote, 2004a) thereby lowering the weight gain in untreated control group. Reduced skeletal growth brought about by mineral deficiency affects growth rate, because skeletal size ultimately determines the capacity of growing animal to accumulate muscle (Sykes et al., 1977). In addition, reduced levels of amino acid incorporation in muscle protein results in reduced weight gains (Soulsby,1982). Reduced weight gain in GI nematode-infected goats was also reported by Howlader et al. (1997) and Githigia et al. (2001). The higher weight gain recorded in tetramisole treated animals is attributable to its higher efficacy against Trichuris spp. and strongyle species when compared to efficacy of albendazole and morantel citrate against these parasites (Jayraw and Raote, 2004b). The present finding is in agreement with Sakhawat et al. (1997), who also observed that levamisole treated sheep gained more weight than albendazole and morantel treated animals. The mean weight gain of 1.97 kg recorded in the present study corroborates well with the findings of Sathianesan and Peter (1972), who recorded 1.7 kg weight gain in tetramisole treated goats over 30 days of observation. The 8.78% weight gain observed in albendazole-treated animals is in general agreement with Faizal et al. (1999) and Githigia et al. (2001). Rajangam and Balchandran (1989) earlier reported a mean weight gain of 38.3 g day-1 in morantel citrate treated goats. Correspondingly, a mean daily weight gain of 55 g day-1 in morantel treated group was also evident in the present study. Das et al. (2004) recorded the per head loss of Rs. 351 and 377 in 3-6 and 6-12 months old GI II III IV 2 3 4 21.05 ±3.09 21.36 ±2.96 21.73 ±3.07 21.57 ±2.4 22.79 ±2.07 21.56 ±3.45 II week 22.09 ±3.16 21.95 ±2.4 23.26 ±1.89 22.05 ±3.44 III week 22.54 ±2.97 22.51 ±2.18 23.84 ±2.21 21.95 ±3.21 IV week 1.82 1.56 1.97 0.79 Mean weight gain (kg) 8.78 7.44 9.00 3.73 Percent weight gain 65 55 70 28 218 187 236 94 44 49 30 - 174 138 206 94 Gross profit Cost of Net profit Mean (Rs.) following medication (Rs.) daily chemotherapy following weight (except Gr.-I) chemotherapy gain (except Gr.-I) (g day-1) Gr.-I: infected untreated control; Gr.-II: treated with tetramisole hydrochloride; Gr.-III: treated with morantel citrate and G r.-IV: treated with albendazole 20.72 ±3.08 20.95 ±2.83 21.87 ±1.85 22.25 ±1.89 21.38 ±3.7 I 1 21.18 ±4.01 I week Sl. Group 0 day No. Table I. Effect of anthelmintic medication on live weight gain in natural GI nematode infection in goats 66 Jayraw and Raote Effect of antihelmenthic therapy on live weight gain nematode-infected Bengal goats, respectively, over a period of nine months. Our findings are also consistent with the findings of above workers, as a net gain of Rs. 206 was recoded in tetramisole treated group followed by Rs. 174 and 138 in albendazole and morantel citrate treated groups, respectively. However, the net profit recorded in infected untreated control group was only Rs.94, which may be attributed to the worm burden, resulting in weight loss. The effective elimination of GI nematodes in all treated groups has a direct bearing on the improved feed utilization ratio and gain in body weight by preventing blood, as well as, protein loss into the GI tract. Tetramisole hydrochloride appears to be superior to albendazole and morantel citrate in effective elimination of GI nematodes and subsequent improvement in weight gain. 67 goats in Kenya. Small Ruminant Research 42: 21-29. Howlader M M R, Capitan S S, Eduardo S L, Roxas N P and Sevilla C C.1997. Performance of growing goats experimentally infected with stomach worm (Haemonchus contortus). Asian Australasian Journal of Animal Sciences 10: 534-539. Jayraw A K and Raote Y V. 2004a Effect of anthelmintic treatment against gastrointestinal nematodes with a note on haematology and plasma proteins in goats. Journal of Veterinary Parasitology 18: 51-54. Jayraw A K and Raote Y V. 2004b Comparative efficacy of tetramisole hydrochloride, morantel citrate and albendazole against gastrointestinal nematodes in goats. Journal of Veterinary Parasitology 18: 131-134. ACKNOWLEDGEMENTS Rajangam R K and Balchandran S. 1989. Efficacy of morantel citrate (Banminth- Pfizer) against gastrointestinal parasites and its effect on body weight gain in stall-fed goats. Indian Veterinary Journal 66: 919 -922. The authors are thankful to the Associate Dean, College of Veterinary and Animal sciences, Parbhani and Farm Superintendent for providing necessary facilities to do this work. Sakhawat A, Anwar A H, Hayat B, Zafar I and Hayat C S. 1997. Field evaluation of anthelmintic efficacy of levamisole, albendazole, ivermectin and morantel tartrate against gastrointestinal nematodes of sheep. Pakistan Veterinary Journal 17: 114-116. REFERENCES Sanyal P K. 1996. Gastrointestinal parasites and small ruminant production in India. Sustainable parasite control in small ruminants. Australian Center of International Agriculture Research Proceedings Series 74: 109-112. Abraham J. 2001. Processing of meat for enhancing its quality and value. Towards Food Security in India, 88th session ISCA, New Delhi. Chowdhury N. 1994. Helminths of domesticated animals in Indian subcontinent. In: Helminthology. Chowdhury. N and Tada. I (Edt.) Narosa and Springer, New Delhi, pp 73-120. Das R, Datta S and Ghosh JD. 2004. Economic impact of gastrointestinal nematodosis in Bengal goat. Fifteenth National Congress of Indian Association for the Advancement of Veterinary Parasitology, Pantnagar, Uttaranchal, India, 25 - 28 October. Faizal A C M, Rajapakse R P V J, Jayasinghe S R and Rupasinghe V. 1999. Prevalence of Eimeria spp. and gastrointestinal nematodes versus weight gains in treated goats raised in the dry areas of Sri Lanka. Small Ruminant Research 34: 21-25. Githigia S M, Thamsborg S M, Munyua W K and Maingi N. 2001. Impact of gastroinestinal helminths on production in Sathianesan V and Peter C T. 1972. Comparative efficacy of thiabendazole, Nilverm and Promintic against gastrointestinal nematodes of goats. Kerala Journal of Veterinary Sciences 3:143-157. Snedecor G W and Cochran W G. 1994. Statistical Methods. 8th ed., Iowa State University Press, Ames, Iowa, pp 135-170. Soulsby E J L. 1982. Helminths, Arthropods and Protozoa of Domesticated Animals. 7th Edition, ELBS and Bailliere Tindall, London, pp 239 - 240. Sykes A R, Coop R L and Angus K W. 1977. The influence of chronic Ostertagia circumcincta infection in the skeleton of growing sheep. Journal of Comparative Pathology 87:521529. Journal of Parasitic Diseases: June 2006, Vol. 30, No. 1, 68–75 J PD Ultrastructure, differential density and distribution pattern of polymorphic microtriches in tegument of Stilesia globipunctata infecting Ovis aries (sheep) C. Venkatesh, K. Ramalingam and V. Vijayalakshmi P.G. and Research Department of Zoology, Govt. Arts College, Chennai. ABSTRACT. Scanning electron and transmission electron microscopic study of tegument of Stilesia globipunctata revealed a complex pattern of microtrichial brush border. They show a wide range of morphological variations. S. globipunctata shows species specific pattern of papilliform, filiform, spiniform and blade-like microtriches. The scolex shows the presence of papilla-like, spine-like and blade-like microtriches. The immature proglottid region is densely covered by posteriorly directed filiform, spiniform and papilliform microtriches. In mature proglottid region, papilla-like and filament-like microtriches are predominantly seen. The microtriches appear to be non-uniform in density and size. In gravid segments, there is obviously an increasing degree of disorder in the arrangement of microtriches. It increases the absorptive surface. It helps to resist the peristaltic movement of intestine and maintains the parasite in position. Its absence in the gravid region is due to the morphological changes in the tegument and the interaction of luminal environment. Keywords: density, distribution, microtriches, polymorphism, Stilesia globipunctata INTRODUCTION The external surface organization of the cestode parasite not only confers protection to the parasite, but also functions as a metabolically active tissue taking up nutrients from the luminal environment and as an outlet for elimination of its extracellular secretions and the excretion phenomenon. It has been noticed that in cestodes, the basic structure may be compared with a gut turned inside out, with the external body covering serving the absorptive functions normally associated with the intestinal mucosa (Smyth and McManus, 1989). Corresponding author: Dr. V. Vijaylakshmi, P. G. and Research Department of Zoology, Govt. Arts college, Men (Autonomous), Nandanam, Chennai - 600 035, Tamil Nadu, India. Ultrastructural studies have shown that the external body covering of the adult tapeworm consists of a naked cytoplasmic layer. There is no resistant cuticle. This outer layer is a syncytium with no partitions of cell walls. It is referred to as a tegument distinguishing it from non-living cuticles of the type that occur in nematode species. Before discussing the functional morphology of the tegument, it is essential to have some understanding of the structural properties of absorptive surfaces in general. The structure, physiology and biochemistry of the tegument are, therefore, of fundamental importance in understanding cestode physiology as a whole. In cestodes, the tegument grows, not by cell division, but by recruitment. Cells form the parenchyma come up and attach to the tegument and vesicles pass up into the tegument. However, the amplification of the 69 Ultrastructure of microtriches in Stilesia globipunctata surface is achieved by the presence of delicate cytoplasmic extensions called microtriches, reminiscent of mucosal cell microvilli. The microtriches on the tapeworm surface increase the surface area of the parasite by about 20-times. The most dominant feature of the cestode tegument is the covering by microtriches, which are thought to be responsible for nutrition and protection and possibly also the mechanical functions of anchoring and traction. They show a wide renge of morphlogical variations. Microtriches are unique in the cestodes and are evident on the tegument and other epithelia that open into the tegument. Microtriches have been widely repeated among all major orders of Eucestoda. These are present on the scolices and strobila of representatives of most groups that have been examined with SEM. Papilliform, spiniform and filiform structures have been variously reported among the Pseudophyllidea (Anderson, 1975, Granath et al., 1983) Cyclophyllidea (Berger and Mettrick, 1971; Ubelaker et al., 1973; Sampathkumar, 2001; Vijayalakshmi, 2001; Radha, 2003). In the present study the characterization of microtrichial structure, density, distribution and their functional significance in S. globipunctata has been attempted as the species of cestode has so far not been exclusively studied due to its inconspicuousness in the gut of sheep. MATERIALS AND METHODS Collection of tapeworms: The tapeworm S. globipunctata (Rivolta, 1874) were collected from the intestine of naturally infacted sheep autopsied in the slaughterhouse at Perambur, Chennai. The sheep intestines were transported to the laboratory within half an hour of collection. In the laboratory, each intestine was carefully dissected and the tapeworms were collected. Then the worms were washed in distilled water to render them free from intestinal contents and rinsed quickly 3-4 times in normal saline. The tapeworms were then observed through a compound microscope to confirm their taxonomic characters. The entire worm was spread out on a board and the length was measured.The immature, mature and gravid proglottid region of the worm was located and separated as follows and dried on moist blotting paper and used for scanning and transmission electron microscopic studies. a. Immature proglottides containing scolex and anterior region. b. Mature proglottides with functional reproductive organs. c. Gravid proglottides containing eggs. Scanning electron microscopic study : The scanning electron microscopic (SEM) studies of the soclex, immature, mature and gravid proglottides of S. globipunctata were undertaken to understand its ultrastructure. For this purpose, the specimens were dissected in chilled glutaraldehyde (2.5%) and fixed for 16 h at 4° C. The tissues were subsequently washed thrice at an interval of 15 min each in phosphate buffer at pH 7.0 and then dehydrated by passing through an ascending series of alcohol from 30 to 100% an h in each concentration. They were ultimately kept in 100% alcohol overnight. Following dehydration, the tissues were air-dried in a desiccator for 7 to 10 days. The dried samples were mounted on an aluminium stub and gold sputtered in vacuum for 10 min, using an Eiko IB-2 ion coater. The samples were observed eventually on a Hitachi, S-415A scanning electron microscope, scanned at 25 KV and micrographed at different magnifications (Hayat, 1977). Transmission electron microscopic study: The scolex, mature and gravid proglottid regions of S. globipunctata for transmission electron microscopy (TEM) were immersed in 2.5% glutaraldehyde in Millong's phosphate buffer (pH 7.3, 380 mOsm/L), where they were diced into small pieces. After 3-4h fixation at room temperature, the tissue was rinsed in Millong's buffer. The tissue was then post-fixed in 1% osmium tetroxide in Millong's buffer for 1.5 h, rinsed quickly in distilled weter, dehydrated in an ethanol series, infiltrated with propylene oxide, embedded in Spurr's low-viscosity epoxy resin and polymerized at 60° C. Thin sections were cut at 70-90 nm with a diamond knife, mounted on uncoated copper grids, stained with uranyl acetate/ethanol and aqueous lead citrate, and examined under a Philips 204 TEM at an accelerating voltage of 40 or 60 kV (Conn, 1993). RESULTS SEM observations: Specimens of S. globipunctata were found to have species-specific patterns of papilliform, filiform, spiniform and blade-like microtriches that are restricted to particular regions of scolex and strobila. The central to peripheral regions of the scolex are covered with papilliform and filiform 70 Venkatesh, Ramalingam and Vijaylakshmi structures. The adherent surfaces of the sucker and their cavities have a dense uniformly distributed covering of spiniform and blade-like microtriches of consistent structure (Fig. 1a). The tegument on the margins and outer surface of the sucker is densely covered with relatively long blade-like microtriches. Similarly, different types of microtriches can also be seen in neck and strobila. The microtriches seem to be directed posteriorly. Microtriches in the center of scolex measure about 2.0 µm and in the sucker region, it measures 2.1µm. The immature proglottid region is also densely covered by posteriorly directed filiform, spiniform and papilliform microtriches (Fig.1b). But the microtriches are not distributed uniformly. This trend continues as segments become older. The polymorphic nature of the microtriches increases as the segments become older. The mature proglottid regions of S.globipunctata clearly reveal the decrease in density and non-uniformity of microtriches (Fig.1c). On gravid segments, however, there is obviously an increasing degree of disorder in the arrangement of microtriches (Fig.1d). The surface of the posterior most part in the majority of worms examined is in a stage of dissolution with on microtriches being apparent. Thus the scanning electron micrographs of the microtriches and their distribution in the tegument of scolex, immature and mature strobilar regions of the parasite revealed the polymorphic nature of the microtriches. Mt Sb Mt (a) (b) Tf Mt Mt (c) (d) Fig. 1. Scanning electron micrographs of scolex, immature, mature and gravid regions showing microtriches of S. globipunctata. a. Adherent surfaces of sucker showing spiniform and blade-like microtriches (x80). b. Tegumental surface of the immature region showing filiform, spiniform and papilliform microtriches (x15). c. Mature region showing posteriorly directed microtriches (with less density; x25). d. Microtriches of gravid region showing non-uniform and less density (x15). Mt - Microtriches, Sb - segental boundary, Tf - tegumental folding. 71 Ultrastructure of microtriches in Stilesia globipunctata TEM observations: In addition to the light and SEM findings, ultrastructural observations were made by TEM. The TEM picture of scolex (Fig. 2a-c) shows the presence of papilla-like, spine-like and blade-like microtriches. The microtriches may be divided into three anatomical regions (Fig. 2b) viz. 1) microfilament containing a base, 2) a dense cap and 3) a complex junctional region between the base and cap. Each base is found to contain an inner sleeve of dense material, the core tunic. Distally, the core is found connected individually to slightly curved tubule, the junctional tubule. As observed in SEM pictures, posteriorly directed filament-like, spine-like and papilla-like microtriches can be seen in the immature proglottid region (Fig. 2d). Whereas in the mature proglottid region, papillalike and filament-like microtriches are predominantly seen (Fig. 3a). The microtriches appear to be nonuniform in density and size. A decreased microtrichial density down the length of strobila and morphological changes in the tegumental surface of the gravid segments can be clearly observed (Fig. 3b). This picture clearly reveals the stages of disintegration of microtriches. Such changes involve surface sculpturing accompanied by loss of all microtriches and erosion of folds in the posterior region of the parasite (Fig. 3c). The disintegrated microtriches can also be seen in Fig. 3d. Cp Cr Cp Jr Jr B Gx Spl B (a) Gx Ct Spl (b) Bm Mt Mt Gx Spl (c) (d) Fig. 2. Transmission electron micrographs of tegument brush border of scolex and immature regions of S. globipunctata. a. T.S. of the tegument showing different kinds of microtriches (x45,000). b. Higher magnification of architecture of blade like mictothrix (x1,000,000). c. L.S. of the margin of suckers showing microtriches (x7000). d. Brush border of tegumental folds of immature region showing different types of microtriches (x20,000). B - Base, Bm - basement membrance, Cp - cap, Ct - core tunic, Gx- glycocalyx, Jr - junctional region, Mt - microtriches, spl - sub plasmalemmal layer. 72 Venkatesh, Ramalingam and Vijaylakshmi Bm Spl Spl (a) (b) Db Mt Spl (c) (d) Fig. 3. Transmission electron micrographs of tegument brush border of mature and gravid regions of S. globipunctata. a. Tegument under higher magnification of mature proglottid region showing different kinds of microtriches (x30,000). b. Tegument (T.S.) of gravid region showing the dissolution of microtriches (x20,000). c. L.S. of tegument of the gravid region showing smooth margin with complete lack of microtriches (x20,000). d. Transmission electron micrographs showing disintegrated microtriches (x70,000). Bm - basement membrane, Db - dense bodies, Mt - microtriches, Spl - subplasmalemmal layer. DISCUSSION The highly active outer surface of cestodes viz. the tegument is a multifunctional entity serving for absorption, digestion, protection, excretion (Featherston, 1972; Thompson et al., 1980; Hayunga, 1991), anchoring (Rothman, 1963; Morseth, 1966; Thompson et al., 1980) and traction for locomotion (Rothman, 1963; 1966; Berger and Mattrick, 1971). At the parasite host interface, it additionally serves for chemical and tactile reception (Featherston, 1972; Webb and Devey, 1974; Jones, 1988; Hass and Guggenheim, 1977; Granth et al., 1983). Although the tegument contains specific systems for the transport of molecules and ions, especially the amino acids, hexose sugars, vitamins, purnes, pyrimidines, nucleotides and lipids, it also serves a number of other vital functions as suggested by Podesta (1980): (a) it is a major site of catalytic activity and contains enzymes of parasite and possibly of host origin, (b) it may be a site of volume regulation, (c) it serves a protective function both against the host's digestive enzymes and the host's immune reactions, (d) it may also function as a site of matabolic transfer (Posdesta, 1982; Pappas, 1983; Threadgold, 1984). Such diverse functions necessitate a high degree of morphological specialization. Jones (1998) has described three features for cestode epithelia, namely, the occurrence of regional specialisation, microtriches and secretory components. In addition he has also suggested that some cestode epithelia are involved in Ultrastructure of microtriches in Stilesia globipunctata developmental processes such as the nurture of embroyos and maintnance of surrounding tissues. The microtriches of cestodes are more complex than mammalian microvilli. An excellent model of their structure is given by Holy and Oaks (1986). TEM studies have, however, revealed that all cestode species hitherto examined possess microtriches and that they are probably of universal occurrence (Morseth, 1966; Yamane, 1986). However, there are differences in shape and density of microtriches between larvae and adult worms (Yamane, 1968). Novak and Dowsett (1983) have observed that during asexual reproduction of T. crassiceps the metacestode tegumental microtriches differentiate into at least three morphologically distinct types. Berger and Mettrick (1971) have described the size, shape and number of microtriches in different parts of the worm of three Hymenolepis species. The study of Anderson (1975) shows that in Diphyllobothrium dendritucum and D. ditremum there is a change in the shape and length of microtriches during development from plerocercoid to adult worms. In H. diminuta, microtriches have been quoted as having maximum diameter of 0.14-0.19 µm and maximum length of 0.91.08 µm (Threadgold, 1984). The presence of microtriches in S. globipunctata has been established in the present study by SEM and TEM. These ordered structures have been linked by many authors to mictovilli of mammalian brush border (Read, 1955; Lumsden and Specian, 1980). There is not only morphological but also functional resemblance, as the parasite absorbs nutrients of low molecular weight through its body surface. In addition to the absorptive functions, the parasite epithlium performs the function of body protection (Odland, 1966). This protective function of the tegument is also attributed to a complicated structure as suggested by Jha and Smyth (1969). The higher level of architectural complexity may reflect a more complex level of function for these surface specialisations of cestodes. The comparison of microtriches among different species thus reveals that the ubuquitous nature of microtriches in all cestodes, which have occupied the luminal niche of the lost animals also have evolved structural and functional features of homology in these different species. The above homology of tegumental structure in different cestode species thus reveals that the distribution microtriches and their further 73 polymorphic modification confers not only advantages to the parasitic species, but also represents as an adaptation for multiple parasitisms. The host's immunological reactions would have also acted as a limiting mechanism to the multiple parasite loads with differential organization. Although the scolex is generally regarded largely as an organ of attachment, in some cestodes such as Echinococcus sp., may also have a 'placental' function and absorb nutrients directly form the mucosal wall, a condition which occurs in some trematodes (Smyth and Halton, 1983). The root like projection with rootlets increases the nutritional surface. Such a nutritional function has been widely reported by Lumsden (1975a) and Lumsden and Hildreth (1983). The microtriches covering the scolex region revealed a structure different from that of the strobila, a situation presumable related to the topography of the host mucosa. The present study revealed marked difference in the distribution pattern and density of microtrichial structures between scolex, and immature and mature segments. The socolex revealed more dense distribution and uniform distribution of microtriches. The immature proglottid region showed dense and non-uniform distribution of microtriches, whereas in the mature region, the microtriches appeared to be non-unitorf in density and size. A decreased microtrichial density down the length of strobila has been noticed. The scolex has also revealed differential nature of the microtriches. The above microtrichial complexity in the scolex region is of parasitic sognificance as it represents the anchoring region and nodular region in the host tissue. In this context, the species S. globipunctata differs from other cestode specieses, which show a simple anchoring device over the intestinal mucosal region of the host. The deep association of the Stilesia sp. in groups, forming nodular regions in the host mucosal tissue is of parasitic importance. Such group association may not have only adverse consequences to the host but it is also difficult to eliminate such group associations than individual parasites anchoring in the host lumen. Berger and Mettrick (1971) have described polymorphism of microtciehes all along the stobilar length. Braten (1968a) has indicated that microtriches 74 form an almost continuous covering of the worm. Earlier ultrastructural studies of T. hydatigena (Featherston, 1972) have revealed three different types of microthrix each associated with a particular area of the strobila. Jha and Smyth (1971) have examined the rostellum of E. graunlosus and reported, "The microtriches and their branches are curved in various directions to form a criss-cross pattern". The surface of the scolex of Silurotaenia siluri is covered with filiform microtriches and giant spine-like and blase-like microtriches. They are also present on the neck region and posterior margins and internal cavities of the suckers (Scholz et al., 1999). Caira and Ruhnke (1991) noted substantial changes in the pattern and distribution fo microtriches during ontogeny of the scolex in Calliobothrium verticillatum. Vijayalakshmi and Ramalingam (2005) observed filament-like, blade-like and intermediate types of microtriches on the tegument of A. lahorea by using SEM and TEM studies. The present SEM and TEM study also clearly revealed the existence of microtrichial polymorphism all along the strobilar length of S. globipunctata. It revealed a complex pattern of microtrochial brush border showing wide range of morphological variations. Species specific pattern of papilliform, filiform, spiniform and blade- like microtriches were also observed in S. globipunctata. In the light of the observations, results and discussions of previous studies, the results on TEM studies on the tegument of S. globipunctata thus infer that the adhesive and absorptive microtriches of the tegument not only allow the diffusion and intake of various nutrients, micro/trace elements and electrolytes indispensable for the growth of the parasites but afford firm positions inside the intestinal cavity wall against the immune factors of the host which could reject the parasite's holdfast. The above dense distribution of microtriches in the scolex region of S. globipunctata also strengthens the above suggestion. Its absence in the gravid region is due to the morphological changes in the tegument and the interaction of luminal environment. REFERENCES Anderson KI. 1975. Comparison of surface topography of three species of Diphyllobothrium (Cestoda, Pseudophyllidea) by scanning electron microscopy. Int J Parasitol 5:293-300. Berger J and Mettrick DF. 1971. Microtrichial polymorphism among hymenolepid tapeworms as seen by scanning electron microscopy. Trans Am Microsc Soc 90:393-403. Venkatesh, Ramalingam and Vijaylakshmi Braten T. 1968. An electron microscopy study of the tegument and associated structures of the plercercoid of Diphyllobothrium latum (L). Z Parasitenk 30:95-103. Caira JN and Ruhnke TR. 1991. A comparison of soclex morphology between the plerocercoid and the adult of C a l l i o b o t h r i m v e r t i c i l l a t u m ( Te t r a p h y l l i d e a : Onchobothridae). Can J Zool 69:1484-1488. Conn DB. 1993. Ultrastructure of the gravid uterus of Hymenolepis diminuta (Platyhelminthes: Cestoda). J Parasitol 794: 584-590. Featherston DW. 1972. Taenia hydatigena. IV Ultrastructure study of the tegument. Z Parasitend 38:214-232. Granath WO, Lewis JC and Esch GW. 1983. An ultrastructural examination of the scolex and tegument of Bothriocephalus acheilognathi (Cestoda: Pseudophyllidea). Trans Am Microsc Soc 102:240-250. Hayat MA. 1977. Principal and Techniques of Electron Microscopy. Biological applications. Vol. 1-9. New York. Van Nostrand Reinhold. Hayunga EG. 1991. Morphologica adaptations of intestinal helminthes. J Parasitol 77:865-873. Hess E and Guggenheim R. 1977. A study of microtriches and sensory processes of the tetrathyridium of Mesocestoides corti hoeppli, 1925, by transmission and scanning electron microscopy. Z Parasitenk 53:189-199. Holy JH and Oaks JA. 1986. Ultra structure of tegumental microvilli (Microtriches) of Hymenolepis diminuta. Cell Tissue Res 244:457-466. Jha RK and Siyth JD. 1969. Echinococcus granulosus: Ultrastructure of Microtriches. Exptl Parasitol 25:232-24. Jha RK and Siyth JD. 1971. Ultrastructure of the rostellar tegument of Echinococcus granulosus with special reference to biogenesis of mitochondria. Int J Parasitol 169-177. Jones MK. 1998. Structure and siversity of cestode epithelia. Int J Parasitol 28:913-123. Lumsden RD. 1975. Parasitological review: Surface and cytochemistry of parasitic helminthes. Exptl Parasitol 37:267-339. Lumsden RD and Hildreth MB. 1983. The fine structure of adult tapeworms. Biology of the Eucestoda. Vol. 1 pp. 177233. Academic Press London. Lumsden RD and Specian R. 1980. The morphology, histology, and fine structure of the adult stage of cyclophylliden tapeworm Hymenolepis diminuta In: The Biology of Hymenolopis diminuta. 147-280. Academic Press, New York. Morseth DJ. 1966. The fine structure of tegument of adult Echinococcus granulosus, Taenia hydatigena and Taenia pisiformis. J Parasitol 52:1074-1085. Ultrastructure of microtriches in Stilesia globipunctata Novak M and Dowett JA. 1983. Scanning electron microscopy of the metacestode of Taenia crassiceps. Int J Parasitol 13: 383-388. 75 Smyth JD and Mc Manus, DP. 1989, The physiology and biochemistry of cestodes. Cambridge University. Odland GF. 1966. Skin. In Histology (ed. R.O. Greep), pp. 420425, New York, London, Sydney, Toronto: McGraw-Hill Book Company. Thompson RCA, Hayron AR and Sue LPJ. 1980. An ultrastructural study of the microtriches of adults Proteocephalus tidswelli (Cestoda: Proteocephalidae). Z Parasitenk 64: 95-111. Pappas PW. 1983. Host - parasite interface. In: Biology of the Eucestoda. Vol. 2 (ed. Arme. C. and Pappas P.W.) 297-334. Academic Press New York. Threadgold LT. 1984. Parasitic platyhelminthes, In: Biology of the tegument (ed. J. Hereiter - Hahn, Maltolsty, A.G. and Richards, K.S.) 132-191. Springer - Verlag, Berlin. Podesta RB. 1980. Concepts of membrance biology in Hymenolepis diminuta. In Biology of the tapeworm hymenolepis, ed. H.P. Arai, pp. 505-509. Academic Press: New York. Ubelaker JE Allison VF and Specian RD. 1973. Surface topography of Hymenolepis diminuta by scanning electron microscopy. J Parasitol 59: 667-671. Podesta RB. 1982. Adaptive features of the surface epithelial wyncytium favouring survival in an immunologically hostile environment. In Parasites-their world and ours, ed. D.F. Mettrick and S.S. Desser, pp. 149-155. Elsevier Biomedical Press: Amsterdam. Radha, T. 2003. Studies on the ultrastructure, biochemical and metabolic aspecits of Raillietina echinobothrida (Megnin, 1881): (Davaineidae), cestode parasite of the fowl host (Gallus domesticus) in relation to host-parasite interaction. Ph.D. Thesis, University Press: New Jersey. Rivolta 1874. Referred from The Fauna of British India Including Ceylon and Burma. Cestoda. Vol. II by Southwell, T. 1930. Rothman AH. 1963. Electron microscope studies of tapeworms. The surface structure of Hymenolepis diminuta. Trans Am Micros Soc 82: 22-30. Rothman AH. 1966. Ultra-structural studies of enzyme activity in the cestode cuticle. Exptl Parasitol 19: 332-338. Scholz T, Zdarska Z, de Chambrier A and Drabek R. 1999. Scolex morphology of cestode Silurotaenia siluri (Batsch, 1786) (Proteocephalidea; Gangesiinae) a parasite of European wells (Silurus glanis). Parasitol Res 85: 1-6. Smyth JD and Halton DW. 1983. The physiology of trematodes, 2nd edn. Cambridge University Press: Cambridge. Sampathkumar. 2001. Studies on the implications of metabolic and biochemical parameters of the larval growth and differentiations of Cysticercus tenuicollis (Cestoda) inside the sheep host (intermediate host). Ph. D., Thesis. University of Madras. Vijayalakshmi V. 2001. Studies on the significance of ultrastructure, bichemical and metabolic parameters in proglottization oriented growth and differentiation of Avitellina lahorea (Woodland, 1927): (Anoplocephalid cestode) inside the sheep host (final host). Ph. D., Thesis. University of Madras. Vijayalakshmi V and Ramalingam K. 2005. Regional difference, ultrastructure and adaptive significance of tegumentary microtrichial polymorphism in Avitellina lahorea (Anoplocephalid cestode) infecting sheep's (Ovis aries) intestine. J Parasitic Dis 29 (1): 9-16. Webb RA and Davey KG. 1974. Ciliated sensory receptors of the unactivated metacestode of Hymenolepis diminuta and H. microstoma. Tissue cell 6: 587-598. Yamane Y. 1968. On the fine structure of Diphyllobothrium eriacei with special reference to the tegument. Yanago Acta Med 12: 169-181. Journal of Parasitic Diseases: June 2006, Vol. 30, No. 1, 76–80 J PD The protozoan fauna living in the digestive system of Periplaneta americana in Kolkata, West Bengal, India J. Ghosh and A. Gayen Post Graduate Department of Zoology, Maulana Azad College, Kolkata. ABSTRACT. The protozoan fauna living in the digestive system of Periplaneta americana in Kolkata, West Bengal, India, was studied. Two ciliates, one amoeba and one mycetozoan were recognized. Of these, Nyctotherus ovalis was the most common and prevalent species (frequency index 92.31, concentration index 4.79). The other ciliate was Balantidium (frequency index 35.55, concentration index 2.57) whose species designation was not very clear. This first study demonstrates the presence of Endamoeba blattae from the hindgut (frequency index 60, concentration index 2.75) and of Coelosporidium periplanetae from Malpighian tubules (frequency index 14.71, concentration index 2.8) that are new records for the protozoan fauna of Kolkata. Additionally, this study highlights various morphological details as well as population distribution pattern of N. ovalis. Keywords: distribution, gut, morphology, Periplaneta americana, protozoa INTRODUCTION Cockroach is a household insect that acts as a mechanical carrier as well as vector to a large number of pathogens. They harbour a variety of protozoans in their digestive system wherein most of them live as endocommensals. The major groups of protozoans reported in the lumen of the gut of cockroaches are ciliates, amoebas, flagellates and apicomplexans (Kudo, 1926a; Kudo, 1926b; Kudo, 1926c; Kudo and Meglitsch, 1938; Hoyte, 1961). Researchers have examined various aspects of parasites including morphological, cytological and ecological details. Roth and Willis (1957) discussed the importance of cockroaches as vectors of various vertebrate pathogens. In the recent past, the contribution(s) of anaerobic protozoans and methanogens to hindgut metabolic activities of American cockroach has been Corresponding author: Mrs. Jayati Ghosh, Post Graduate Department of Zoology, Maulana Azad College, Kolkata-700 013, W.B., India. E-mail: jayati1973@yahoo.com studied in detail (Gijzen and Barugahare, 1992). Surprisingly, in India, especially in West Bengal, Kolkata, only a few studies of this kind have been reported. In 1922, two new ciliates Balantidium ovatum and B. blattarum (Ghosh, 1922a; Ghosh, 1922b) were reported. In a description of Indian ciliophorans, Nyctotherus ovalis was reported to be a part of protozoan fauna of Periplaneta americana (Bhatia and Gulati, 1927). But afterwards, apparently, no detailed studies were undertaken on this aspect. Therefore, presently, it is difficult to draw a list of the fauna of cockroach gut for these areas without making proper investigations. We, therefore, considered it expedient to study the fauna of the digestive system of P. americana in Kolkata. Such studies can be expected to generate a new list of species, and the data related to their prevalence (number of host infected) and intensity (average number of parasites/host). The goal of present investigation, therefore, was to study the protozoan fauna of P. americana in Kolkata, and to determine their morphology and distribution. Our findings suggest that N. ovalis is the most 77 Protozoan parasites of Periplaneta americana common and dominant specimen, which showed considerable variation in its nuclear structure. Large numbers of N. ovalis were observed in the colon of P. americana with only a few specimens of Balantidium, but its species status was not very clear because the organisms had some morphological similarities and dissimilarities with Balantidium blattarum. This is the first study wherein we report the presence of Endamoeba blattae and Coelosporidium periplanetae in considerable numbers in the hindgut and Malpighian tubules, respectively, of P. americana collected from West Bengal, India. Nevertheless, these two protozoan species have already been reported earlier in P. americana from some other countries (Kudo, 1926a, Sprague, 1940). MATERIALS AND METHODS The cockroaches used in this study were collected from Beliaghata, East Kolkata households and Belgachia market area during June 2004 to February 2006. The cockroaches were dissected within 24 h of their collection. The mid-gut, ileum, colon and rectum of P. americana were then taken out separately and kept in different watch-glasses containing saline (0.6% NaCl in distilled water) solution. The population counts were made through haemocytometer following Petroff-Hauser counting method (Prescott et al., 1999). The gut smears were first examined under a light microscope and then permanent preparations were made. Fixation was done by Schaudinns' fixative and the smears were stained with iron-alum haematoxylin (Cable, 1963). For the study of nuclear characters, Feulgen nuclear technique (Feulgen and Rossenbeck, 1924) was utilized. An ocular micrometer calibrated with an objective micrometer was used for all the cell measurements. All measurements given in the study are in micrometers, and the figures were drawn by using a camera lucida. RESULTS AND DISCUSSION Data in Table I show the population distribution pattern of the parasites obtained from the gut of P. americana. A comparative population distribution pattern of the parasite species showed that mean density range of N. ovalis was highest (4.79) followed by C. periplanetae (2.8), E. blattae (2.75) and Balantidium sp. (2.57). These data corroborate with the previously published results by Gijzen and Barugahare in 1992. But the maximum SD value was attained by N. ovalis (2.83) and the lowest was observed in E. blattae (1.55). Table I: Comparative population distribution pattern of protozoan fauna in the hindgut of P. americana Name of specimen Number of host studied Number of host infected Frequency index Density range Mean Standard deviation (SD) Standard error (SE) N. ovalis Balantidium sp. E. blattae C. periplanetae 26 90 40 34 24 32 24 5 92.31 35.55 60 14.71 (1-12) x 104 (1-7) x 104 (1-5) x 104 (1-6) x104 4.79 2.57 2.75 2.8 2.83 1.73 1.55 1.72 0.58 0.31 0.32 0.77 Table II: Data of various morphological characters of N. ovalis Body length (L) Body width (W) L/W Macronucleus length (ML) Macronucleus width (MW) ML/MW No. of Specimens Range (in µm) Mean (in µm) Standard deviation (SD) Standard drror (SE) 25 25 25 25 25 25 57-123.5 47.5-95 1.13-1.8 19-47.5 9.5-23.75 1-3 87.21 62.32 1.43 25.46 14.63 1.923 11.93 12.83 0.20 6.97 4.64 0.56 2.39 2.57 0.04 1.39 0.93 0.11 78 Ghosh and Gayen Description of the specimens: stains lightly. Also variable forms of macronucleus are common in N. ovalis. Available forms here are spherical, ovoid, cylindrical, club shaped, semi-ovoid forms. Light micrograph of N. ovalis showing variation in nuclear shape (Fig. 2). N. ovalis Leidy, 1849 Habitat: Maximum number of specimens was found in the colon of hind-gut. Genus Balantidium Claparède and Lachmann,1858 Morphology: Various morphological characteristics of N. ovalis have been studied including body length (L) and width (W) and their ratios (L/W), macronucleus length (ML) and width (MW) and their ratios (ML/MW; Table II). Habitat: Colon of the hind-gut. Morphology: Body was pear shaped, anterior end tapering, slightly bent to the side opposite to the peristome. Small peristome observed about onefourth the body length. Endoplasm was coarsely granular. Macronucleus was spherical (diameter 9.514.25 µm) and placed behind the peristome in the center of the body. Body length varied from 57-76 µm and width from 38-57 µm. Fig. 3 shows general morphology of Balantidium sp. under a light , microscope, stained with Heidenhain s iron hematoxylin. A large contractile vacuole was present at the posterior end in B. blattarum (Ghosh, 1922b) that was lacking in this specimen. The body observed to be oval, comparatively wider in posterior than anterior. Elongated macronucleus situated at anterior one-third. Body length varied from 57-123.5 µm and width from 47-95 µm. Cytopyge terminal and oval or slit like in shape. Peristome begins at the anterior end turns slightly to the right and ends in cytostome located midway between the ends. Nuclear length varied from 19-47.5 µm and width from 9.5-23.75 µm. In our study, body dimensions of N. ovalis were relatively smaller than those described by Leidy, 1849. When the L/W ratio was considered, it can be said that the specimens were slightly oval whereas ML/MW ratio indicated that nuclear shape varied from spherical to ovoid and elongated. E. blattae Bütschli, 1878 Habitat: Colon and ileum of hind-gut. Morphology: Amoeba with few, broad pseudopodia and showed distinct ectoplasm and endoplasm. Endoplasm was clear and homogeneous. Nucleus with characteristic round or ovoid shape of this parasite can be easily distinguished. Dark granules are also found along its peripheral region with light Fig. 1 shows the general morphology of N. ovalis under a light microscope and stained with Heidenhain,s iron hematoxylin (a) and using Feulgen nuclear stain (b). Compact macronucleus gave strong positive reaction in Feulgen test where micronucleus An V Ma Ma CP P An Cy 9.5 µm 9.5 µm P (a) (b) Fig. 1. General morphology of N. ovalis under a light microscope: (a) stained with Heidenhain,s iron hematoxylin, (b) using Feulgen nuclear stain. An. anterior, P. posterior, V. vestibular opening, CP. cytopharynx, Ma. macronucleus, Cy. cytopyge. 79 Protozoan parasites of Periplaneta americana An An 19 µm An An Ma V Ma Ma Cy Ma V CP 19 µm P 19 µm 19 µm P (a) P P (c) (b) Cy (d) Fig. 2. Light micrograph of N. ovalis showing variation in nuclear shape: (a) ovoid, (b) club-shaped, (c) cylindrical , (d) semi-ovoid. An. anterior, P. posterior, V. vestibular opening, CP. cytopharynx, Ma. macronucleus, Cy. cytopyge. central part. Length of the amoeba varies from 66.5152 µm and width 47.5-104.5 µm. Nuclear diameter was about 19-47.5 µm. Fig. 4 shows general morphology of E. blattae under a light microscope following staining with Heidenhain , s iron hematoxylin. C. periplanetae Lutz and Splendore, 1903 19 µm PS N Habitat: Malpighian tubule. Morphology: Trophozoite stages are common with developing spore inside. Shape of the trophozoite may be spherical, ovoid or cylindrical. Diameter of spherical trophozoites varies from 28-31.5 µm and in En Ec An Fig. 4. General morphology of E. blattae under a light microscope, stained with Heidenhain,s iron hematoxylin. Ps. pseudopodia, N. nucleus, En. endoplasm, Ec. ectoplasm. V Mi Ma 19 µm P Fig. 3. General morphology of Balantidium sp. under a light microscope, stained with Heidenhain,s iron hematoxylin. An. anterior, P. osterior, V. vestibular opening, Mi. micronucleus, Ma macronucleus. case of ovoid ones observed length is 42 µm and width 35 µm. Fig. 5(a), (b) and (c) show light micrograph of C. periplanetae stained with Heidenhain,s iron hematoxylin showing trophozoites containing various stages of developing spore. The above findings indicate that the fauna of P. americana collected from Kolkata is quite rich in various species of protozoan parasites. For the last fifteen years, ciliates in the cockroach gut have received special emphasis due to the discovery of hydrogenosome, a double-membrane sub-cellular organelle, present in some anaerobic protist including N. ovalis (Müller, 1993). Biochemical and molecular genetic evidences argue that hydrogenosome share a common ancestry with mitochondria (Embley et al., 1997). However, all hydrogenosomes studied so far 80 Ghosh and Gayen 30 µm 30 µm CB CB CB T DS (a) 30 µm (b) (c) , Fig. 5. Light micrograph of C. periplanetae stained with Heidenhain s iron hematoxylin showing trophozoites containing various stages of a developing spore: (a), (b) and (c), CB. chromatoid bodies, T. trophozoite, DS. developing spore. lack a genome. Recently Akhmanova et al. (1998) isolated hydrogenosomal DNA from N. ovalis. Hydrogenosomes in N. ovalis are intimately associated with endosymbiotic, methanogenic bacteria that may have a major role in cockroach metabolism. In vitro studies also suggest a major role for hind-gut protozoa in cockroach metabolic activities, especially during the insect growth period (Gijzen and Barugahare, 1992). ACKNOWLEDGEMENTS The authors would like to acknowledge University Grants Commission, New Delhi, for providing financial support for this work [Minor project No. PSW/032/04-05(ERO)]. The authors would also like to thank Prof. Dipankar Sengupta, Head, Dept. of Zoology, Maulana Azad College, Kolkata, for providing departmental facilities and to other colleagues for continuous cooperation. The authors are especially thankful to Prof. Biswapati Dasgupta and Dr. Tarak Khan for their valuable suggestions and comments. REFERENCES Akhmanova A, Voncken F, van Alen T, van Hoek A, Boxma B, Vogels G, Veenhuis M and Hackstein JHP. 1998. A hydrogenosome with a genome. Nature 396:527-528. Feulgen R and Rossenbeck H. 1924. Mikroskopischchemischer Nachweis einer Nucleinsarure von Typus der Thymonucleinsaure und die darauf beruhende elective Farbung von Zellkernen in mikroskopischen Praparaten. Ztschr. Physiol. Chem. 135:203. Ghosh E. 1922a. On a new ciliate, Balantidium ovatum sp. nov., an intestinal parasite in the common cockroach (Blatta americana). Parasitology 14:371. Ghosh E. 1922b. On a new ciliate, Balantidium blattarum sp. nov., an intestinal parasite in the common cockroach (Blatta americana). Parasitology14:15-16. Gijzen HJ and Barugahare M. 1992. Contribution of anaerobic protozoa and methanogens to hindgut metabolic activities of the American cockroach, Periplaneta americana. Appl. Environ. Microbiol. 58: 2565-2570. Hoyte HMD. 1961. The protozoa occurring in the hind-gut of cockroaches I. Parasitology 51:415-436. Kudo R.1926a. Observations on Endamoeba blattae. Am. J. Hyg. 6: 139-52. Kudo R.1926b. Observations on Lophomonas blattarum, a flagellate inhabiting the colon of the cockroach, Blatta orientalis. Arch. Protistenk. 53: 191-214. Kudo R. 1926c. A cytological study of Lophomonas striata Bütschli. Arch. Protistenk. 55: 504-515. Kudo RR and Meglitsch PA.1938. On Balantidium praenucleatum n.sp. inhabiting the colon of Blatta orientalis. Arch. Protistenk. 91:111-124. Müller M. 1993. The hydrogenosome. J. Gen. Microbiol. 139:2879-2889. Bhatia BL and Gulati AN. 1927. On some parasitic ciliates from Indian frogs, toads, earthworms and cockroaches. Arch. Protistenk. 57:85-120. Prescott LM, Harley JP and Klein DA. 1999. Microbiology.4th Edn. WCB/McGraw-Hill, USA. pp 117. Cable RM. 1963. An illustrated laboratory manual of parasitology, Allied Pacific Private Limited, Bombay, India, pp133-134. Roth LM and Willis ER. 1957. The medical and veterinary importance of cockroaches. Smithson. misc. Collns.134: no. 10 Embley TM, Horner DA and Hirt RP. 1997. Anaerobic eukaryote evolution: hydrogenosomes as biochemically modified mitochondria? Trends Ecol. Evol. 12: 437-441. Sprague V. 1940. Observations on Coelosporidium periplanetae with special reference to the development of the spore. Trans. Am. Microsc. Soc. 59: 460-474. Journal of Parasitic Diseases: June 2006, Vol. 30, No. 1, 81–84 J PD Chelatrema neilgherriensis n. sp. (Trematoda: Gorgoderidae) infecting the freshwater fishes from Noolpuzha river in Wynad district, Kerala, India K. T. Manjula and K. P. Janardanan Department of Zoology, University of Calicut, Calicut. ABSTRACT. A new species of digenetic trematode Chelatrema neilgherriensis (Gorgoderidae) was recovered from the freshwater fishes Danio neilgherriensis and Labeo rohita, collected from Noolpuzha river in Wynad district, Kerala. Herein, we have described this new species in detail, discussed its systematic position and have compared it with the only other species in the genus C. smythi Gupta and Kumari, 1973. We found C. neilgherriensis n. sp. to be different from C. smythi in body measurements, size of oral and ventral suckers, sucker ratio, extent of uterine coils, nature of testes and seminal vesicle, and shape and size of eggs. Apparently, this first study reports a second species of the genus Chelatrema, a gorgoderid from Kerala. Keywords: Chelatrema, Gorgoderidae, Kerala, trematoda INTRODUCTION The genus Chelatrema was erected by Gupta and Kumari (1973) with C. smythi as the type species which infect the freshwater fish, Chela bacala (Ham.) from Satluj River at Ropar, Punjab, India. The authors included the genus under the subfamily Arnolinae Yamaguti, 1958 of the family Hemiuridae Luhe, 1901. But Gibson and Bray (1979) stated that it is not a hemiurid, and Gibson (2002) considered it as a gorgoderid. During an explorative study on the trematode fauna of freshwater fishes in the hill streams of Wynad district in Kerala, we came across a species of Chelatrema in Danio neilgherriensis and Labeo rohita, collected from Noolpuzha river. Detailed studies proved that the trematode is significantly different from C. smythi, Corresponding author: Dr. K. P. Janardanan, Department of Zoology, University of Calicut, Calicut-673 635, Kerala, India. E-mail: kpjanardanan@yahoo.co.in the only species in the genus Chelatrema, and is reported here as a new species. This forms the second species of the genus Chelatrema and the first report of a gorgoderid from Kerala. MATERIALS AND METHODS The fresh water fishes Danio neilgherriensis (Day, n=16) and Labeo rohita (Hamilton, n=6), collected from Noolpuzha river in Wynad district of Kerala, from April 2004 to May 2005, were examined for trematode infections. The flukes recovered from the intestine of D. neilgherriensis and the intestine and body cavity of L. rohita were observed under phasecontrast microscope, with or without vital staining. Those used for permanent preparations were fixed for approximately 1 h in 10% formalin under cover glass pressure at room temperature, and stained with alum carmine as described by Cantwell (1981). The descriptions are based on 20 whole mounts, and measurements of 10 mature flukes fixed in 10% formalin. Measurements are presented in 82 Manjula and Janardanan micrometres, unless otherwise stated; the range is followed by mean values in parentheses. The figures were drawn with the aid of a camera lucida, and details added free-hand from observations made on live specimens. RESULTS Infections with adult flukes were observed in the intestine of 10 out of 16 (62.5%) Danio neilgherriensis, and intestine and body cavity of one out of six (16.7%) Labeo rohita. The intensity of infection varied from one to three. Description (Fig. 1) Body large, light red, elongate, ovoid, narrowing in anterior body; measured 2.7146.5 x 1.0112.602 (4.413 x 1.747) mm; width 30-45 (40)% of body-length. Tegument unarmed. Oral sucker round, sub-terminal; 390-625 (490) in diameter. Ventral sucker round, muscular, large; 510-1,035 (801) in diameter. Sucker length ratio 1: 1.69-1.82 (1.78). Anterior body 9002,192 (1,962) long; 30-37 (34)% of body length. Prepharynx distinct, 20-72 (64) in length. Pharynx elongate, ovoid; measured 125-232 x 94-215 (163 x 136). Oral sucker/pharynx width ratio 1: 0.30-0.38 (0.34). Oesophagus 18-42 (29) long. Many gland cells found scattered around oesophagus and anterior region of caeca. Intestinal bifurcation in anterior body; 800-1,205 (1,016) anterior to ventral sucker. Caeca narrow, 3.0-4.985 (3.872) mm long; terminate blindly, 320-456 (389) from posterior extremity. Testes two, symmetrical, spherical to ovoid, entire; left testis 250-432, 209-382 (311 x 246); right testis 249-382 x 197-309 (305 x 243). Post-testicular region 820-1,198 (1,006) long; 18-27 (22)% of body-length. Vas deferens traverses along left side of ventral sucker. 75 µm 800 µm B A 30 µm 300 µm C D Fig. 1. A-D. C. neilgherriensis n. sp. A, Entire worm. B, Terminal genitalia. C, Egg. D, Miracidium. 83 A new species of Chelatrema from fishes in Wynad Cirrus-sac large, elongate, post-bifurcal, medially placed; 279-415 x 107-193 (366 x 159) in size. Seminal vesicle saccular, bipartite; 212-384 x 51-97 (348 x 77), reaches about ¾th of cirrus-sac, narrows distally forming pars-prostatica. Pars-prostatica thick-walled, ensheathed in numerous gland cells. Ejaculatory duct long, tubular, with narrow walls; opens into base of genital atrium. Cirrus not visible. Genital pore submedian, post-bifurcal; 162-283 (246) from ventral sucker. Ovary immediately posterior to ventral sucker, equatorial, dextral, round to ovoid; 301-439 x 376-483 (345 x 406). Ventral sucker to ovary distance 84-129 (113); 2-3.1 (2.5)% of body-length. Oviduct leads from lateral margin of ovary. Seminal receptacle large, saccular, ovoid, placed lateral to ovary; 300-452 x 223-327 (361 x 255) in size. Vitellarium in the form of a compact mass, situated behind seminal receptacle, on the left side of median line; 180-362 x 150-286 (276 x 225) in size. Uterus fills the whole post-testicular posterior body, extends into entire extra-caecal space, up to the level of caecal bifurcation. Distal end of uterus passes along left side of ventral sucker to form distinct metraterm and opens at genital pore. Uterus filled with numerous, small, oval, thin-shelled, nonoperculate, embryonate, and fragile eggs, measuring 78-86 x 70-81 (83 x 76). 232 (1,309 x 201), extends to midlevel of testes. Taxonomic summary Genus: Chelatrema Gupta and Kumari, 1973 Species: Chelatrema neilgherriensis n. sp. Type host: Danio neilgherriensis (Day), Cyprinidae Additional host: L a b e o ro h i t a ( H a m i l t o n ) , Cyprinidae Site: Intestine and body cavity Type locality: India, Kerala, Wynad district, Noolpuzha river Holotype: Deposited in the Department of Zoology, University of Calicut, Kerala, India. No: Z./Par./Dig2005-1a Paratypes: Z./Par./Dig-2005- lb-d Date of collection: 13 April 2005 Etymology: Named after the species name of the type host. Excretory bladder long, I-shaped; 1,190-1,570 x 104Table I. Comparison of C. neilgherriensis n. sp. with C. smythi Gupta and Kumari, 1973 Character C. smythi Gupta and Kumari, 1973 C. neilgherriensis n. sp. 1. Body size 6.9-7.0 x 2.5-2.75 mm 2.714-6.5 x 1.011-2.602 (4.413 x 1.747) mm 2. Oral sucker Round; 600 x 500-550 Round; 390-625 (490) 3. Ventral sucker Round; 1,000 x 900 Round; 510-1,035 (803) 4. Sucker length ratio 1 : 1.6 1 : 1.78 5. Testes Entire or slightly lobed Entire, round to ovoid 6. Seminal vesicle Ovoid; 66 x 88 Bipartite; 212-384 x 51-97 (348 x 77) 7. Ovary Entire; 500 x 400 Entire, round to ovoid; 301-439 x 376-483 (345 x 406) 8. Uterine coils Fill the space behind ventral sucker Fill the space behind ventral sucker, and extend extra-caecally up to the level of caecal bifurcation. 9. Eggs Small, round; 45-60 Small, ovoid; 78-86 x 40-49 (83 x 46) C. bacala (Hamilton) D. neilgherriensis (Day) and L. rohita (Hamilton) 10. Host (s) Note: Measurements are in micrometres unless otherwise mentioned. 84 Miracidium Freshly laid eggs hatched in 2-3 h. Miracidia pyriform, with conical papilla at anterior end; posterior end round; measured 132-176 x 46-73 (151 x 64). Cilia long, uniformly distributed. Eyespots two, oval and conjoined; 19-23 x 8-13 (20 x 11.5) in size. A large apical gland and a pair of lateral penetration glands present. A pair of flame cells present behind the eyespots. Miracidia swim actively in water, changing direction intermittently. They exhibited negative phototaxis. DISCUSSION In the present study, the fluke is characterised by the presence of large ventral sucker, well-developed pharynx, short oesophagus, symmetrical testes, pretesticular ovary, single compact vitellarium, strongly convoluted uterus and embryonated eggs, and hence it is included in the genus Chelatrema Gupta and Kumari, 1973. These authors included the genus under family Hemiuridae Luhe, 1901, but Gibson and Bray (1979) treated it as a genus inquirendum and Gibson in 2002 considered it a gorgoderid. As the characters of the genus Chelatrema agree fully with that of Gorgoderidae, we agree with the arrangement made by Gibson and place the genus under the family Gorgoderidae. The new species reported here needed to be compared with the only species in the genus, C. smythi which infects the freshwater fish Chela bacala. It resembled C. smythi in the extent of caeca, position of gonads, nature and position of vitellarium and position of genital pore. But it differed greatly in body measurements, size of oral and ventral suckers, sucker Manjula and Janardanan ratio, extent of uterine coils which lead extra-caecally up to caecal bifurcation, nature of testes and seminal vesicle and the shape and size of eggs. A comparison of the characters of C. smythi and of C. neilgherriensis, presented in Table I, showed that the two species were different from each other. Further, they infect different hosts too. Therefore, the fluke reported herein has been considered as a new species and named Chelatrema neilgherriensis after the specific name of the type host, Danio neilgherriensis. ACKNOWLEDGEMENTS The authors express their sincere thanks to Dr. D. I. Gibson, Department of Zoology, The Natural History Museum, London, for taxonomic identification of the fluke to generic level, and to the Head of the Department of Zoology, University of Calicut, for providing facilities. REFERENCES Cantwell GE. 1981. Methods for invertebrates. In: staining procedures. Clark G (Ed.) Williams and Wilkins, Baltimore. pp 255-280. Gibson DI. 2002. Family Derogenidae Nicoll, 1910. In: Keys to the Trematoda. Volume 1. Gibson DI, Jones A & Bray RA (Eds.) CAB International. pp 351-368. Gibson DI and Bray RA. 1979. The Hemiuroidea: terminology, systematics and evolution. Bull. Br. Nat. Hist. (Zool.). 36: 35-146. Gupta N K and Kumari A. 1973. Chelatrema smythi n. gen. n. sp. (Trematoda, Hemiuidae: Arnolinae) from a fresh-water fish, Chela bacala, from Ropar. Res. Bull. (N.S.) of the Panjab Univ. 24:109-112. Short communication Journal of Parasitic Diseases: June 2006, Vol. 30, No. 1, 85–88 J PD Haemato-biochemical studies on fowl coccidiosis in layer birds* N. D. Hirani, J. J. Hasnani, R. S. Joshiand K. S. Prajapati Department of Veterinary Parasitology, College of Veterinary Science and Animal Husbandry, Anand Agricultural University, Anand. ABSTRACT. This study on haemato-biochemical profile of 48 coccidia-infected and 24 healthy birds from commercial layer farms revealed that the infected-birds had significantly (p < 0.05) lower mean haemoglobin concentration (7.62 ± 0.14 vs 10.59 ± 0.19 g%) and total erythrocyte count (1.63 ± 0.04 vs 3.10 ± 0.05 million cells/cmm), apparently lower packed cell volume (23.00 ± 0.71 vs 31.46 ± 0.70 unit %) and significantly (p < 0.05) higher mean total leucocyte count (16.41 ± 0.37 vs 10.40 ± 0.16 thousand cells/cmm) than the healthy ones. The mean values of blood glucose (221.01 ± 2.66 vs 176.03 ± 0.75 mg%) and serum total cholesterol (271.71 ± 4.71 vs 112.68 ± 0.60 mg%) were significantly (p < 0.05) higher and serum total protein was lower (2.98 ± 0.08 vs 4.42 ± 0.10 g%) in coccidia-infected birds as compared to healthy birds, whereas the activities of serum aspartate aminotransferase (77.84 ± 1.10 vs 65.62 ± 0.63 U/L), alanine aminotransferase (9.20 ± 0.28 vs 7.07 ± 0.25 U/L) and alkaline phosphatase (902.23 ± 9.30 vs 816.58 ± 25.95 KAU%) were non-significantly higher in infected-birds than the healthy ones. Almost a similar trend was observed for all the parameters in layer birds managed under both deep litter as well as cage system. These findings reflected anaemic condition with depressed metabolism due to tissue damage and increased immune response in infected-birds. Keywords: coccidiosis, deep litter vs cage system, haematology, layer birds, serum Coccidiosis is a parasitic disorder, which can have an acute or chronic course of infection in birds. It produces a deviation in the various haematological, biochemical and enzymatic components of the body (Padmavathi and Muralidharan, 1986a,b). Therefore, measurement of biochemical and enzyme activities is useful in determining the pathologic conditions in the tissues. Serological profiles of certain enzymes are altered greatly in cell membrane degeneration, and inflammatory and diffuse tissue * A part of M. V. Sc. thesis of the first author; approved by A A V, Anand. Corresponding author: Dr. N. D. Hirani, Department of Veterinary Parasitology, College of Veterinary Science and Animal Husbandry, Anand Agricultural University, Anand-388 001, Gujarat, India. degeneration and loss (Deger et al., 2002). Scanty literature is available on haemato-biochemical alterations due to coccidiosis in layer birds from round the globe. This study was, therefore, conducted on 99 layer farms in the middle of Gujarat, involving fecal sampling of 594 birds managed under deep litter and/or cage system. All the samples were examined by the standard methods. From all coccidia positive farms, two infected and one healthy birds were selected for haemato-biochemical studies. In all, 48 blood samples from coccidia-infected birds and 24 samples from normal healthy birds were studied. Data thus generated were statistically analyzed using unpaired 't' test (Snedecor and Cochran, 1980). In this study, significantly (p < 0.05) lower mean 86 haemoglobin concentration and total erythrocyte count, apparently lower packed cell volume, and significantly (p < 0.05) higher mean total leucocyte count were found in coccidia-infected birds than the healthy ones (Table I). These findings were in conformity with the earlier reports (Joshi et al., 1974; Padmavathi and Muralidharan, 1986a; Jaipurkar et al., 2004). The reduction in the value of haemoglobin and total erythrocyte count observed in the infected-birds might be attributed to haemorrahges in the intestine followed by development of intestinal lesions. There may be injury to tissue and liberation of large quantities of histamine, which increase the local blood flow and also increase the permeability of capillaries and venules allowing large quantities of fluid to come out (Padmavathi and Muralidharan, 1986a). The reduction observed in packed cell volume during the acute-phase of infection might be due to severe blood loss resulting in anaemic condition (Joshi et al., 1974). The increased total leucocyte count in coccidiainfected birds might be due to immune suppression of infection. This was suggestive of increased leucopoiesis as a first step of defense mechanism to infection (Padmavathi and Muralidharan, 1986a). In the present study, significantly (p < 0.05) higher blood glucose and serum total cholesterol and lower serum total proteins were found in coccidia-infected birds as compared to healthy ones. Joshi et al. (1974) reported similar findings on blood glucose. However, Padmavathi and Muralidharan (1986b) recorded serum hypoglycaemia in birds with experimental Emeria tenella infection, whereas Basith et al. (1998) failed to see any change in plasma glucose. This high level of blood glucose observed in coccidia-infected birds may be either due to stress conditions leading to the liberation of adrenal corticoids which induce hyperglycaemia or disturbed carbohydrate metabolism due to interference with phosphorylative carbohydrate dissimilation by unidentified material present in the intestine of infected-fowls. The apparent hyperglycaemia was undoubtedly due to loss of erythrocytes. The observations on protein were in conformity with those of Turk (1972), who reported marked reduction in protein during the acute-phase of E. necatrix infection. The significant (p < 0.05) reduction in serum total protein observed in infectedbirds might be due to reduced feed intake and/or haemorrahges through the gut and formation of inflammatory exudates rich in blood proteins (Basith et al., 1998). The decrease might also be due to leakage of protein resulting from the increased permeability of Hirani et al. the intestinal mucosa, which was necessary for the passage of plasma protein at the height of the disease. These lower protein levels might be due to reduced absorption of amino acids derived from the protein constituents of feed due to reduced feed consumption (Padmavathi and Muralidharan, 1986b). The results on cholesterol are in agreement with the observations of Singh et al. (1976), Padmavathi and Muralidharan (1986b) and Basith et al. (1998), whereas Constantinescu (1976) could not see significant changes in serum cholesterol values in both infected and healthy birds. The hypercholesteremia observed in the present study among the infected-birds might be due to disturbed fat metabolism and loss of fluid resulting in apparent increase (Padmavathi and Muralitharan, 1986b) or due to impaired liver function consequent to injury to intestinal epithelium in coccidiosis (Basith et al., 1998). Mean activities of serum aspartate aminotransferase, alanine aminotransferase and alkaline phosphatase observed in infected-birds were non-significantly higher as compared to the healthy ones. Singh et al. (1976) reported comparable findings, while Kumar and Rawat (1975) found no significant difference, but values were slightly less in infected group as compared to normal group. The increase in SGOT and SGPT activities observed in coccidia-infected birds under study might be due to extensive damage to intestine and liver by the parasites and thereby loss of appetite. Constantinescu (1976) reported significant increase in serum AKP activity among infected-birds. On the contrary, Kumar and Rawat (1975) reported significant decrease in serum AKP of E. necatrix- and E. acervulina-infected 3-4 months old cockerels. It can be concluded that haemoconcentration, reduced haemoglobin and total erythrocyte count and an increase in total leucocyte count in coccidia-infected birds suggest haemorrhage and increased leucopoiesis as a first step of defense mechanism to infection. The high level of blood glucose observed in coccidiainfected birds may be either due to stress leading to the liberation of adrenal corticoids which induce hyperglycaemia concomitant with loss of erythrocytes. Reduced serum total protein and increased serum enzyme activity in coccidia-infected birds indicate damage to intestine and vital organs, increased cell membrane permeability and loss of intracellular contents. 1.70 ± 0.08 15.33 ± 0.94 230.53 ± 6.15 3.22 ± 0.20 278.55 ± 7.11 76.13 ± 2.61 9.49 ± 0.53 Erythrocytes count (million cells/cmm) Leukocytes count (thousand cells/cmm) Blood glucose (mg %) Serum total protein (gm %) Serum cholesterol (mg %) SGOT (U/L) SGPT (U/L) N = number of birds , * Significant at p < 0.05, 884.58 ± 15.79 24.68 ± 1.69 Packed cell volume (%) Alkaline phosphatase (KAU %) 7.87 ± 0.39 12 Infected birds Haemoglobin (gm %) N Parameter 834.91 ± 29.41 7.18 ± 0.32 65.63 ± 0.69 112.43 ± 0.59 4.48 ± 0.13 175.21 ± 0.74 10.47 ± 0.20 3.03 ± 0.04 31.06 ± 0.77 10.48 ± 0.24 18 ns = non-significant. 908.11 ± 11.18 9.10 ± 0.33 78.41 ± 1.19 269.43 ± 5.82 2.90 ± 0.08 217.84 ± 2.74 16.77 ± 0.37 1.61 ± 0.05 22.44 ± 0.76 7.54 ± 0.13 36 Healthy birds Cage system Infected birds ** Significant at p < 0.01, 761.57 ± 52.57 6.77 ± 0.19 65.58 ± 1.52 113.40 ± 1.70 4.26 ± 0.14 178.50 ± 1.70 10.20 ± 0.17 3.30 ± 0.11 32.67 ± 1.61 10.93 ± 0.16 6 Healthy birds Deep litter 902.23 ± 9.30 9.20 ± 0.28 77.84 ± 1.10 271.71 ± 4.71 2.98 ± 0.08 221.01 ± 2.66 16.41 ± 0.37 1.63 ± 0.04 23.00 ± 0.71 7.62 ± 0.14 48 Infected birds 816.58 ± 25.95 7.07 ± 0.25 65.602 ± 0.63 112.68 ± 0.60 4.42 ± 0.10 176.03 ± 0.75 10.40 ± 0.16 3.10 ± 0.05 31.46 ± 0.70 10.59 ± 0.19 24 Healthy birds Overall total Table I. Haemato-biochemical profile of coccidia-infected and healthy layer birds managed under deep litter and cage system in Gujarat 0.60ns 0.93ns 1.49ns 4.85** 1.93* 2.40* 2.24* 3.79** 1.41ns 2.22* 't' value Haemato-biochemical studies on fowl coccidiosis 87 88 ACKNOWLEDGEMENTS We thank the Principal and Dean of the College, and the poultry farmers of the region for the facilities, support and co-operation extended for this work on their birds. The facility of autoanalyzer provided by ADIO, Navsari, Gujarat State is also gratefully acknowledged. Hirani et al. Joshi HC, Singh BP, Prasad B and Prasad RK. 1974. Variations in certain blood constituents during caecal coccidiosis in poultry. Indian J Parasitol. 9:22-24. Kumar A and Rawat JS. 1975. A note on the effect of coccidiosis on serum enzymes, blood glucose and cholesterol in chicken. Indian J Anim Sci 45:154-156. REFERENCES Padmavathi P and Muralidharan SRG. 1986a. Alteration in haematological parameters in chicken during Eimeria tenella infection. Indian Vet J 63:716-722. Basith Abdul S, Rajavelu G and Murali Manohar B. 1998. Biochemical studies in experimental Eimeria necatrix infection in chickens. Indian Vet J 75:876-878. Padmavathi P and Muralidharan SRG. 1986b. Studies on the alteration in the serum metabolites during the Eimeria tenella infection in chicks. Indian Vet J 63:530-536. Constantinescu V. 1976. Biochemical and histoenzymic changes in coccidiosis in chickens. Buletinul Institutului Agronomic Cluj Napoca 30:115-117. Singh CV, Joshi HC and Shah HI. 1976. Biochemical studies in intestinal coccidiosis of poultry. Pantnagar J Res 1:63-66. Deger Y, Dedo S and Deger S. 2002. Enzyme activity changes in the sera of chickens treated with coccidiostatic agents. Indian Vet J 79:912-916. Jaipurkar SG, Deshpande PD, Narladkar BW, Rajurkar SR and Kulkarni GB. 2004. Caecal coccidiosis in broiler chicks: haematological, pathological changes during treatment with herbal antidiarrhoels. J Vet Parasitol 18:135-138. Snedecor GW and Cochran WG. 1980. Statistical Methods. 8th Edn. Iowa State Univ. Press, Ames, Iowa, USA. Turk DE. 1972. Protozoan parasitic infections of the chick intestine and protein digestion and absorption. J Nutr 102:1217-1222. Journal of Parasitic Diseases: June 2006, Vol. 30, No. 1, 89–91 Short communication J PD Re-redescription of Dissurus farrukhabadi Verma, 1936 (Digenea - Echinostomatidae) with a discussion of the genus Dissurus Verma, 1936 P. C. Gupta and R. B. Singh Department of Zoology, D. A. V. College, Kanpur. ABSTRACT. Dissurus farrukhabadi Verma, 1936 from the large intestine of a White-necked stork, Dissurus episcopus (Boddaert) from Unnao, India is re-redescribed. The validity of the species of Dissurus, Verma, 1936 is discussed. D. xenorhynchi Wahid, 1962 is considered a synonym of Stephonoprora gigantica Gupta, 1962 and Psilocollaris guptai Gupta and Saxena, 1986 is considered a synonym of D. farrukhabadi. During the survey of digenetic termatode parasites of birds, nine specimens of Dissurus farrukhabadi Verma, 1936 were collected and described. The trematodes were fixed in 70% alcohol under slight cover glass pressure for 24 h, stained with acetic alum carmine, dehydrated in graded series of alcohols, cleared in clove oil and mounted in Canada balsam. The drawings were made with the aid of a camera lucida, and unless otherwise stated, all measurements are in mm. DESCRIPTION Dissurus farrukhabadi Verma, 1936 Body long, slender, delicate, spinose, 5.34-5.71 x 0.57-0.69, with rounded extremities. Head collar reniform, 0.30-0.34 x 0.41-0.45; with 24 spines, interrupted dorsally, arranged in a single row with no end group. Oral sucker terminal, avoid 0.10-0.12 x 0.08-0.09. Prepharynx 0.06-0.09 long. Pharynx subglobular 0.11-0.13 x 0.09-0.16. Oesophagus tubular, with thick irregularly sinuous walls, 0.48Corresponding author: Dr. P.C. Gupta, 8/200-A, Arya Nagar, Kanpur-208 002, U. P., India. 0.58 long. Intestinal caeca simple, extending to hind end of body. Ventral sucker sub-spherical, much larger than oral sucker, 0.40-0.46 x 0.43-0.50 at 0.87-1.14 from anterior extremity. Excretory bladder Y-shapped; arms bifurcation just behind posterior testis and their extension beyond anterior testis not traceable due to vitallariaum follicles, excretory pore subterminal. Testes oval, entire or notched, subequal, tandem, apart, in last fifth of body. Anterior testis 0.30-0.55 x 0.23-0.28 at 3.61-4.30 from anterior extremity. Posterior testis, 0.47-0.60 x 0.31-0.32 at 0.48-0.52 from posterior extremity. Vasa efferentia arising from testis, join together to form vas deferens, running anteriorly to form vesicula seminalis. Cirrus sac small, heart shaped, 0.21-0.27 x 0.14-0.17, extending posteriorly either upto anterior margin of ventral sucker or overlapping anterior third of it. Vesicula seminalis bipartiate, 0.12-0.15 x 0.09-0.16. Parsprostatica small, 0.03-0.06 long surrounded by large number of prostate gland cells. Cirrus muscular, 0.010.02 long. Genital pore median, between intestinal bifurcation and ventral sucker at 0.72-0.99 from anterior extremity. 90 Gupta and Singh Ovary rounded, entire, submedian, postequatorial, pretesticular, intercaecal, 0.14-0.18 x 0.16-0.18 at 2.73-3.22 from anterior extremity. Receptaculam seminalis oval, 0.044 x 0.035. Laurer's canal present. Vitellaria follicular, dense extending from a level anterior to ovary up to hind end of body. Transverse vitelline ducts from either side meet and form a yolk reservoir to open at ootype, surrounded by a large number of Mehlis' gland cells. Uterus long, intercaecal lying between ootype and venteral sucker, Egg large, oval, few, 0.09-0.10 x 0.04-0.05. Host : White-necked stork Dissurus episcopus (Bodd.) Location : Large intestine Locality : Unnao, U.P., India Material : Nine specimens (incidence 2/3) Verma (1936) erected the genus Dissurus with D. farrukhabadi as its type, recovered form the intestine of a White-necked stork. D. espicopus from Farrukhabad, U.P. Wahid (1962) added D. xenorhynchi from the large intestine of Black-necked stork. Xenorhynchus asiaticus at London zoo, and Dwivedi (1967) described D. setheae from the type host at Chindwara (M. P.). Srivastava (1974) reexamined Verma's specimens and gave illustrations together with an emended generic diagnosis. Singh (1954) established another genus Psilocollaris with P. indicus as its type for his specimens from the intestine of D. episcopus from Lucknow, U.P. The genus Psilocollaris was characterized by an aspinous collar. Gupta (1980) critically studied P. indicus Singh, 1954 in detail and raised question on its validity, doubting its conspecificity with D. farrukhabadi. Srivastava 2 LC 0.25mm MG RC 3 0.05mm 4 0.2mm 5 6 0.05mm 1 Fig. 1-6. Dissurus farrukhabadi Verma, 1936. 1. entire ventral view; 2. head collar, (enlarged); 3. extension of cirrus sac up to anterior third of ventral sucker; 4. cirrus sac, enlarged; 5. female genital complex, enlarged; 6. showing entire testes. OV - ovary, U - uterus, OO-ootype, RS - receptaculum seminalis, LC - Leurar's canal, YR - yolk reservoir. Re-redescription of Dissurus Farrukhabadi Verma (1982) re-examined the type specimen of P. indicus and found the presence of collar spines and accepted the views of Gupta (1980). Further, he considered P. singhi Pandey, 1975 as species inquirenda and also transferred D. setheae to the genus Stephanoprora Odhner, 1902, due to presence of 22 collar spines, tandam testes lying in mid-third of body, vitellaria extending form posterior testis to hind end and short uterus. Srivastava (1982) further observed that D. setheae is probably a synonym of S. nigerica Gupta, 1983. The authors are in agreement with the views of Gupta (1980) and Srivastava (1982). In the opinion of present authors D. xenorhynchi Wahid, 1962 better fits under the genus Stephanoprora Odhner, 1902 and shares most of its characters viz. the extension of the vitellaria, 22 collar spines, the position of the testes and thickening in the wall of the oesophagus with S. gigantica Gupta (1962). The former, therefore, is considered a synonym of later. Gupta and Saxena (1986), overlooking the synonymy of Psilocollaris, described P. guptai form the intestine of an Indian stork, Leptoptilos dubius form Lucknow and differentiated it from the type species on such characters as the muscular pharynx, position of the testes and the ovary. These characters are merely intraspecific variations and, therefore, P. guptai is a synonym of D. farrukhabadi. The present specimens of D. farrukhabadi differed from the previous descriptions in having a smaller body, in the extensions of vitellaria from a little anterior to overy instead of from the level of it, and in having a smaller post-testicular space. These differences are considered as intraspecific variations. REFERENCES Dwivedi MP. 1967. A new species of the genus Dissurus Verma, 1936 (Trematoda Echinostomatidae). Natural and Applied Science Bulletin, 20: 267-275. 91 Gupta PD. 1980. Further observations on Psilocolaris indicus Singh, 1954 with a note on its systematic position (Trematoda: Psilostomatidae). Bulletin of the Zoological Survey of India. 2: 217-218. Gupta R. 1962. Studies of trematode parasites of Indian birds. II. On Stephanoprora gigantics sp. nov. from the Blacknecked storck, Xenorhynchus asiaticus (Latham). Proceeding of the National Academy of science, India. 32:381-386. Gupta R. 1963. On Stephanoprora nigerica sp. nov. with a brief review of the genus Stephanoprora Odhner, 1905 (Trematoda : Echinostomatidae). Zoological Anazica. 170:117-130. Gupta V and Saxena AM. 1986. On Psilocollaris guptai sp. nov. (Psilostomatidae : Trematoda) from the intestine of an Indian Stork, Leptonhilos dubius (Gmelin) from Lucknow. Indian Journal of Helmonthlogy. 37: 1-3. Pandey KC. 1975. Studies on some known and unknown trematode parasites. Indian Journal of Zootomy. 14: 197219. Singh KS. 1954. Psilocollaris indicus n.g., n. sp. (Psilostomatidae) from an Indian stork, Dissurus episcopus episcopus. Journal of the Washington Academy society. 44: 24-26. Srivastava CB. 1974. A critical study of Verma's "Notes on trematode parasites of Indian birds" based on his trematode collection on Part 2. Family Echinostomatidae Dietz, 1909. Journal of the Zoological society of India. 24:160-191. Srivastava CB. 1982. The fauna of Indian and the adjacent countries. Platyhelminthes Vol. I (Supplement) Trematoda Digenea. Addition to Prof. H. R. Mehra's volume on trematoda - Digenea. Publ. by the Director, Zoolgical Survey of India, Calcultta, 163pp. Verma SC. 1936. Notes on trematode parasites of Indian birds. Part I Allababad University Studies. 12: 148-188. Wahid S. 1962. On a new trematode from Black-necked stork, Xenorhynchus asiaticus. Journal of Helminthology. 36: 211-214. Short communication Journal of Parasitic Diseases: June 2006, Vol. 30, No. 1, 92–93 J PD A case of vaginal bleeding due to leech bite R. P. Ganguly, M. S. Mukhopadhyay and K. K. Patra Department of Obstetrics and Gynaecology, R. G. Kar Medical College and Hospital, Kolkata. Medically important leeches belong to Phylum Annelida and Class Hirudinea. It is a worm like creature that attaches to their hosts by means of chitinous cutting jaws and draws blood through muscular suckers. Haemadipsa sylvastris, H. zelanica and H. montana, H. ornata etc. are land leeches available in India. The most prevalent species is H. sylvestris. Hirudo birmanica and Limnatis nilotica are water leeches. L. nilotica is notorious for internal hirudiniasis. Internal hirudiniasis may involve pharynx, larynx, vocal cord, trachea, oesophagus and genitourinary tract. Attachment is usually painless. Continued bleeding from the site takes place after the leech has detached. Death due to exhaustion has been reported. Leech gets dislodged from its site of attachment if saline water, strong vinegar or match flame is applied. CASE REPORT A six year old girl was admitted as an emergency case in the Department of Gynaecology and Obstetrics at R. G. Kar Medical College and Hospital, Kolkata, with a complaint of bleeding per vagina for 4 h. Patient's mother gave history that her daughter had gone to a local pond in the evening to wash herself after defecation. She came back home and after sometime noticed that her undergarments were wet with blood. Patient denied any history of local trauma. She was then brought to the hospital for medical help. On examination, the patient was conscious but very pale. On local examination, fresh blood was seen coming out through vagina. The bleeding point could not be seen as it was inside vagina. There was no Corresponding author: Dr. R. P. Ganguly, BC 45/7, Bichitra Abason, Sector 1, Saltlake, Kolkata-700 064, W.B., India. evidence of trauma anywhere. Per rectal examination revealed a boggy mass of about 5 cm x 4 cm which was appreciated anteriorly. On milking the swelling towards introitus blood clot came out along with a leech and the swelling disappeared. Under intra venous sedation vaginal douching was done with normal saline. The hysteroscope was introduced inside vagina which revealed a bleeding point on the lateral wall of vagina about 1.5 cm from introitus. No other trauma in the vagina could be seen. A paediatric Foley's Catheter (No.10) was introduced inside vagina and the bulb of Foley's Catheter was inflated to 20 cc with distilled water. Three hundred ml (300 ml) of blood was transfused to the patient. Catheter was kept in vagina for 4 h. During the period there was no bleeding from vagina. Routine blood examination and coagulation profile showed normal picture. Patient was kept admitted for 1 week before she was discharged as leech bite patient often gets recurrent episodes of bleeding. This patient too had little trickle on the third day but did not require any intervention . We present this unusual case of vaginal bleeding in a six-year old girl because this is a rare cause of vaginal bleeding and its management by Foley's bulb tamponade avoids unnecessary and complicated intervention under general anaesthesia in a child. It also maintains anatomical integrity of local parts. The leech is a parasite which has a habit of entering anatomical orifice of human being and animals. It releases an anticoagulant (heparin like substance, hirudinin) during the process of sucking blood which causes bleeding from bite site. Deka and Rajeev (2001) have reported leech bite as an unusual cause of haematuria. Severe rectal bleeding from leech bite case of vaginal bleeding was reported by Raj et al. (2001), and vaginal bleeding resulting from leech bite was reported by Hernandez and Ramirez (1998). REFERENCES Deka P M and Rajeev T P. 2001. Unusual cause of haematuria. Urol Int 66:41-42. 93 Hernandez M and Ramirez RE. 1998. Vaginal bleeding resulting from leech bite. Ginecol Obstet Mex 66:246-248. Raj S M, Radzi M and Tee M H. 2000. Severe rectal bleeding due to leech bite. Gastroenterol 95:1607. Short communication Journal of Parasitic Diseases: June 2006, Vol. 30, No. 1, 94–97 J PD Field evaluation of a rapid immunochromatographic test kit for the diagnosis of Plasmodium falciparum and non-falciparum malaria parasites from Sonitpur district, Assam C. Rajendran and S. N. Dube Biotechnology Division, Defence Research Laboratory, Tezpur. ABSTRACT. The sensitivity and specificity of ICT Parascreen test kit device (rapid test for malaria pan/Pf) was compared with conventional microscopic method as the gold standard. A total of 126 patients were tested with ICT and microscopic blood-smear examination. Sixty one (48.41%) patients were found to be positive for malaria infection in microscopic examination. With the ICT kit, the sensitivities and specificities for Plasmodium falciparum and non-falciparum (P. vivax) parasites were 96.30 and 88.88%, and 98.48 and 98.48 %, respectively. The ICT kit gave a little lower sensitivity as compared to microscopic examination. Keywords: HRP II, ICT Parascreen test kit, malaria, pLDH Globally, malaria still remains a major parasitic disease with high morbidity and mortality. In the Northeast region of India, the endemicity of malaria infection and the predominance of Plasmodium falciparum over P. vivax is now well documented (Dev and Phookan, 1996; Kamal and Das, 2001). Accurate diagnosis and proper treatment, which are important to the control of malaria, are the main aims of the global malaria control strategy (WHO, 2000a). Based on clinical symptoms alone, malaria and non-malaria febrile cases and the species of Plasmodium can not be distinguished (Chandramohan et al., 2001). Further, accurate diagnosis of malaria is essential to make a rational choice of drug for the treatment. The development of Rapid Diagnostic Tests (RDT) for malaria based on immunochromatographic test (ICT) strips provides a valid alternative to conventional Corresponding author: Dr. Rajendran, Biotechnology Division, Defence Research Laboratory, Post Bag No. 2, Tezpur-784 001, Assam, India. E-mail: chellairajendran@rediffmail.com microscopic method (WHO, 1996). The ICT is a RDT for P. falciparum specific HRP II antigen (Parra et al., 1991) and a pan malaria antigen (Garcia et al., 1996). RDT is very much useful where microscopy is lacking and where the malaria is severe (WHO, 2000b). The conventional microscopy is the gold standard test to diagnose malaria infections worldwide. Even though it is sensitive and economical, several disadvantages make it inconvenient for use in the field viz., requirement of electricity, trained microscopy personals and difficulties in accurate species identification as evidenced by sensitive molecular methods. Currently, rapid ICT cards are available to detect malaria infection upto species level (Wangsrichanalai, 2001). The first generation of detection kits was designed to diagnose only P. falciparum, but now the newer devices have been designed to diagnose both P. falciparum specific antigens as well as Plasmodium genus specific antigens. Early detection and differentiation of malaria is of paramount importance due to the Immunochromatographic test kit evaluation for malaria diagnosis incidences of cerebral malaria and drug resistance associated with falciparum malaria, and due to the morbidity associated with the other malaria forms. So, ICT kits are considered for malaria diagnosis as microscopic method has limitations. So far, field evaluation of ICT kits has not been done for its use in immediate clinical management of malaria in Sonitpur district, Assam. Therefore, the present study was undertaken to study the performance of the ICT kit in the field by comparing it with the gold standard microscopy method. The study was conducted at seven different places of Sonitpur district, Assam, between August 2004 and May 2005. Each patient, who was suffering from fever, was finger pricked by using a sterile lancet. Thick and thin blood-smears were prepared and stained with Giemsa according to standard procedures. The thick smear was used to detect the infection, whereas the thin smear was used for species identification. A blood sample was considered negative, when no parasite could be detected in 100 fields of an oil immersion (x1000 magnification) objective lens of a microscope (Fernando et al., 2004). At the same time, approximately 5 µl of whole blood from finger prick of the patient was transferred directly to a sample pad. For testing the sample, ICT Parascreen test kit (Zephyr Biomedicals, India) was used as rapid diagnostic device (Lot No: 101003, Mfg Dt: 08-2004 and Exp. Dt: 07-2006). Parascreen utilizes the detection of P. falciparum specific histidine rich protein II, which is water soluble protein that is released from parasitized-erythrocytes of infected individuals, whereas for the detection of pan malaria, Parascreen detects the presence of pan malaria specific pLDH released from the parasitised erythrocytes. Then, a drop of buffer was added and allowed to react for 2 min. This buffer was added to induce cell lysis and allow PfHRP II and pan-malarial antigens to bind to colloidal gold-labeled monoclonal antibodies. All tests were considered as valid if a control line was observed. The sensitivity and specificity was calculated as per the formula given by Mason et al. (2002). The sensitivity was calculated as the number of true positives by the test divided by total positives by Giemsa [TP/(TP+FN)], and the specificity was determined as true negatives divided by the false positives (TN/TN+FP). A total of 126 blood samples were collected. Among these, 61 (48.41 %) samples were found positive for malaria infection by standard microscopical blood- 95 smear examination. Out of these 61 positive samples, 52 were found positive for P. falciparum (85.24 %) and 9 were positive for non-falciparum parasite (P. vivax; 14.75 %), whereas the Parascreen ICT kit showed 54 samples positive for P. falciparum and eight for P. vivax. A summary of the findings by these two tests has been given in Table I. The sensitivity and specificity values of the ICT were calculated using the results of microscopic examination as the gold standard. As per the Parascreen ICT kit, the sensitivity and specificity for P. falciparum and non-falciparum (P. vivax) parasites were 96.30 and 88.88 %, and 98.48 and 98.48 %, respectively. The present field study revealed that the ICT kit could detect P. falciparum from 54 cases, whereas light microscopy showed 52 positive cases. Among the 54 cases, two blood-smears were found negative for P. falciparum but the ICT was positive for them. The possible reason could be due to the persistence of PfHRP II following the clearance of P. falciparum (Wongsrichanalai et al., 1999). But for the case of nonfalciparum (P. vivax) parasite, the ICT kit could detect only eight cases out of nine found by microscopical method. Generally, irrespective of the manufacturers, the sensitivity and specificity for non-falciparum, with the available ICT kit, is varying from 50-70% and 37.580%, respectively, as compared to microscopy. But the present study revealed 88.88% sensitivity and 98.48% specificity. This study was conducted based on qualitative assessment but not quantitative one as the accurate counting of parasites was not carried out to assess the correlation of the sensitivity. But increasing sensitivity of the test with increasing parasite densities is one of the main factors in detecting parasites (Mason et al., 2002). Various authors have reported different degrees of sensitivity and specificity of P.f/P.v test kit manufactured from different countries and from different localities. Palmer et al. (1998) evaluated OptiMAL test for the diagnosis of P. falciparum and P. vivax malaria, and it uses a monoclonal antibody to the intracellular antigen parasite lactate dehydrogenase (pLDH). It differentiates species by the use of a P. falciparum specific and a genus specific antibody. It has shown 88 and 94% sensitivities on symptomatic Honduran patients and specificities of 100 and 99% for the diagnosis of falciparum and vivax malarias. Beatriz E ferro et al. (2002) evaluated OptiMAL and gave a higher efficiency of 98.1% for P. vivax than 94.9% for P. falciparum, in a malaria referral center in Colombia. But Mason et al. (2002) evaluated two test kits viz, 96 OptiMAL and ICT of which both gave lower sensitivities than the earlier report. ICT gave 86.2 and 2.9% sensitivity for P. falciparum and P. vivax and specificity of 76.9 and 100%, but OptiMAL gave 42.6 and 47.1% sensitivity and, 97 and 96.9% specificity for P. falciparum and non-falciparum parasites, respectively. Kolaczinski et al. (2004) evaluated OptiMAL for the diagnosis of P. vivax and P. falciparum with 34% as compared to 36% by gold standard microscopy. For OptiMAL 48 test, cross checking of the corresponding smears at the reference laboratory gave a sensitivity of 79.3% and a specificity of 99.7% for P. falciparum, and the corresponding values of 86.1% and 98.7% for P. vivax infections. The performance of the field microscopy was better, with a sensitivity and specificity of 85.2 and 99.7% for P. falciparum, and 90.4 and 98.7% for P. vivax, respectively. According to Tjitra et al. (1999), the ICT malaria P.f /P.v test was 96% sensitive and 90% specific for P. falciparum, and 75% sensitive and 95% specific for P. vivax even though the blood samples were collected in an anticoagulant, EDTA, from the veins but not from the finger prick. Forney et al. (2000) conducted a study in Thailand and Peru with P.f/P.v test kit and recorded 95% sensitivity and 85% specificity for P. falciparum, and 68% sensitivity and 87% specificity for P. vivax and concluded that the sensitivities were closely correlated with parasite densities. Richter et al. (2004) evaluated ICT (Malaria ICT Now, Binax, Portland, USA) for its performance on 2547 patients, and ICT was positive in all of 204 patients with symptomatic P. falciparum infections, whereas microscopy revealed parasites in 202 of 204 of these patients. But, from these two cases, trophozoites were detected by microscopy only in samples taken after 6 and 12 h. So ICT showed a specificity of 99.74%, whereas for non-falciparum malaria, the sensitivity and specificity of ICT was only 50 and 37.5%, respectively, as compared to microscopy. In our present study, one out of nine cases was found negative for non-falciparum (P. vivax) by ICT. Moreover, this particular blood smear had a comparatively very low density of parasites (2 parasites/100 microscopic fields). Even though the parasite could be seen on blood smear, the ICT missed to detect the infection. Moody et al. (2000) conducted a study and concluded that though parasite may look healthy in the blood smear but it may be non-viable and not producing pLDH. It is known that pLDH activity gets declined with therapy. However, the ICT kit may be a useful device where microscopy is not available, and immediate clinical diagnosis of malaria Rajendran and Dube is required especially for P. falciparum cases which may develop cerebral complications. But for nonfalciparum malaria cases, the present study has given a moderate percentage of sensitivity and specificity but it is difficult to suggest on this aspect, as the number of cases studied was small. With the ICT P.f/P.v the sensitivity for the detection of P. vivax was 96% at parasitaemias greater than 500 parasites/µl blood but, at parasitaemias lower than this the sensitivity was reduced to only 29% (Tjitra et al., 1999; Hunt-Cooke et al., 1999). The ICT can not replace the microscopy method for the determination of parasitaemia. Several factors may be responsible for a little lower performance of the test kit viz, human error though the manufacturer's instructions were strictly followed. The information regarding the history of selfmedication by the patients and the storage condition of the kits also need consideration. Moreover, increasing sensitivity of the test with increasing parasite densities is one of the main factors in detecting parasites at low densities rather than the human error (Mason et al., 2002). So, therefore, to assess the sensitivity and specificity of ICT for non-falciparum parasites, further studies should be done with larger number of samples for the evaluation of its detection limit. Table I. Showing the number of cases studied for the evaluation of ICT with standard microscopic method Parasite Giemsa method ICT (Parascreen) kit P. falciparum 52 54 Non-falciparum (P. vivax) 9 8 Negative 65 - Total 126 - ACKNOWLEDGEMENTS The authors are grateful to The Director, Defence Research Laboratory, Tezpur, Assam, for providing necessary facilities and constant encouragement. The study was not supported by the manufacturer of the ICT kit. REFERENCES Beatriz E Ferro, Iveth J Gonzalez, Fanny de Carvajal, Gloria I Palma and Nancy G Saravia. 2002. Performance of OptiMAL in the diagnosis of Plasmodium vivax and Plasmodium falciparum infections in a malaria referral Immunochromatographic test kit evaluation for malaria diagnosis 97 center in Colombia. Mem Inst Oswaldo Cruz, Rio de Janeiro 97:731-735. immunochromatographic tests to expert microscopy in the diagnosis of malaria. Acta Tropica 82:51-59. Chandramohan D, Carneiro I, Kavishwar A, Brugha R, Desai V and Greenwood B. 2001. A clinical algorithm for the diagnosis of malaria results of an evaluation in an area of low endemicity. Tropical medicine and International Health. 6: 505-510. Moody A. 2002. Rapid diagnostics tests for malaria parasites. Clinical Microbiology Reviews 15:66-78. Dev V and Phookan S. 1996. Malaria prevalence in Tea Estates of Brahmaputra valley of Assam, India. Journal of Parasitic Diseases 20:189-192. Forney J R, Wongsrichanalai C, Magill A J, Sorichaisinthop J and Gasser R A. 2000. Performance characteristics of the ICT Malaria P.f, P.v (TM) rapid diagnostic device for the detection of P.f and P.v. Abstract No: 239, Presented at the 49th Annual Meeting of the American Society of Tropical Medicine and Hygiene, Houston.Tx, USA, Oct 29 Nov 2,2000. American Journal of Tropical Medicine and Hygiene 62:235-236. Fernando D, Karunaweera N D, Fernando W P, Attanayake N and Wickremasinghe A R. 2004. A cost analysis of the use of the rapid, whole blood, immunochromatographic P.f/P.v assay for the diagnosis of P. vivax malaria in a rural area of Srilanka. Annals of Tropical Medicine and Parasitology 98:5-13. Garcia M, Kirimoama S, Marlborough D, Leafasia J and Rickmann K H. 1996. Immunochromatographic test for malaria diagnosis. Lancet 347:1549. Hunt-Cooke A, Chiodini P, Doherty T, Moody A, Reis J and M Pinder.1999. Comparison of parasite lactate dehydrogense based immunochromatographic antigen detection assay (OptiMAL) with microscopy for the detection of malaria parasites in human blood samples. American Journal of Tropical Medicine and Hygiene 60:173-176. Kamal S and Das S C. 2001. Epidemiological observations on malaria in some parts of Darrang district, Assam. Indian Journal of Malariology 38:25-31. Kolaczinski J, Mohammed N, Ali I, Khan N, Ezard N and M Rowland. 2004. Comparison for the detection of Plasmodium vivax and Plasmodium falciparum: Considerations for the application of the rapid test in Afghanistan. Annals of Tropical Medicine and Parasitology 98:15-20. Mason D P, Kawamoto F, Lin K, Laoboonchai A and Wongsrichanalai C. 2002. A comparison of two rapid field Palmer C J, Lindo J F, Klaskala W I, Quesada J A, Kaminsky R, Baum M K and A L Ager. 1998. Evaluation of the OptiMAL test for rapid diagnosis of Plasmodium vivax and Plasmodium falciparum malaria. Journal of Clinical Microbiology 36: 203-206. Parra M, Evans C and Taylor D W. 1991. Identification of P.f histidine rich protein 2 in the plasma of human with malaria. Journal of Clinical Microbiology 26: 1629-1634. Richter J, Harms G, Stover I M, Gobels K and Haussinger D. 2004. Performance of an immunochromatographic test for the rapid diagnosis of malaria. Parasitology Research 92:518-519. Tjitra F, Suprianto S, Dyer M, Currie B J and Anstery N M. 1999. Field evaluation of the ICT Malaria P.f /P.v immunochromatographic test for detection of Plasmodium falciparum and Plasmodium vivax in patients with a presumptive clinical diagnosis of malaria in Eastern Indonesia. Journal of Clinical Microbiology 37:2412-2417. Wangsrichanalai C, Chuanak N, Tulyayon S, Thanoosingha N, Laoboonchai A, Thimasarn K, Brewer TG and Heppner DG. 1999. Comparison of a rapid field immunochromatographic test to expert microscopy for the detection of Plasmodium falciparum asexual parasitemia in Thailand. Acta Tropica. 73: 263-273. Wangsrichanalai C. 2001. Rapid diagnostic techniques for malaria control. Trends in Parasitology (formerly Parasitology Today) 17:307-309. World Health Organization, 1996. A rapid dipstick antigen capture assay for the diagnosis of falciparum malaria. WHO informal consultation on recent advances in diagnostic techniques and vaccines for malaria. Bulletin of the World Health Organization 74:47-54. World Health Organization, 2000a. WHO Expert Committee on Malaria. Twentieth Report. Technical Report Series No.892. Geneva.WHO. World Health Organization, 2000b. Report of a joint WHO/USAID. Informal consultation 25th to 27th October 1999. WHO/CDS/RBM/200.14. Geneva.WHO. J PD JOURNAL OF PARASITIC DISEASES Instructions to Authors General Journal of Parasitic Diseases is the official organ of the Indian Society for Parasitology (ISP). Manuscripts are accepted from international authors irrespective of whether they are members of the ISP or not. The scientific standard as related to quality, originality, clarity and brevity is the sole criterion for accepting a manuscript for publication. Each volume of Journal of Parasitic Diseases consists of two issues released in June and December. “Instructions to Authors” are published in the June issue of the journal each year. Authors are requested to read these instructions very carefully and follow them strictly to ensure efficient and quick editorial processing and publication of their manuscripts. Manuscripts that are incomplete or are not prepared strictly according to the instructions and style of the journal will be returned without editorial processing. Aims and scope The primary constituency of Journal of Parasitic Diseases is parasitology. It publishes original research papers (pure, applied and clinical), which contribute significantly to any area of parasitology. Research papers on various aspects of cellular and molecular parasitology are especially welcome. Submission of manuscripts All manuscripts, in triplicate (three hard copies + 1 disk copy) along with a duly complete “Undertaking by Authors”, and correspondence should be addressed to: Professor Prati Pal Singh, Ph. D., F. N. A. Sc. Editor-in-Chief Journal of Parasitic Diseases Room No. 113, Block-F National Institute of Pharmaceutical Education and Research Sector-67, Phase-X S. A. S. Nagar-160 062, Punjab INDIA E-mail: drppsinghjpd@yahoo.co.in Alternatively, the manuscripts (Word format or PDF files; text, references, tables, legends and figures) and undertaking in electronic form can be submitted as an e-mail attachment. However, simultaneously, they must also be submitted as three hard copies + 1 disk copy along with the hard copy of the “Undertaking by Authors”. Manuscripts are received with the understanding that the research reported therein was conducted with proper animal care as per the approved guidelines of the Institutional Animal Ethics Committee. Similarly, manuscripts which report experiments on human subjects and materials, must have obtained necessary approvals from the relevant institutional ethics committees. Submission of a paper to Journal of Parasitic Diseases implies that all the authors have approved submission of the paper, that the paper is not under consideration for publication elsewhere, and that if the paper is accepted for publication or published, the article or portions thereof will not be published elsewhere without written consent from the Editor-in-Chief. It is the responsibility of the author(s) to obtain all the necessary permission to include items like figures, tables and other materials. In the selection of papers for publication, the decision of the Editor-in-Chief is final and no query regarding the decision will be entertained. Statements or opinions expressed in Journal of Parasitic Diseases reflect the view(s) of the author(s) and do not represent the official position of the ISP. Neither the Society nor the Editorial Board accepts any responsibility for the statements made by the contributors. Forms of publication Journal of Parasitic Diseases publishes contributed papers falling into the following categories. • Full papers: that report original research. • Short communications: those report some preliminary but important findings that do not warrant publication as full paper. • Letters to the Editor-in-Chief • Reviews: including solicited and contributed ones on topics of current importance. Review of manuscripts From now on, the journal shall follow “Tutorial Editorial Policy”. Manuscripts received by the Editorial office, after checking their completeness and suitability as per the scope of the journal, shall be sent to appropriate Members of the Editorial Board, who in turn shall send them to two suitable Reviewers /Referees/ Consultants. They will read, evaluate and return the manuscripts along with their report/comments. The Editorial Board Member, on the basis of the report/comments received and on the scientific quality of manuscript, shall send his or her recommendations (regarding acceptance, nonacceptance or modification) to the Editor-in-Chief. Finally, the Editor-in-Chief, based on all the reports and recommendations and on the overall merit of the manuscript, shall decide and communicate the corresponding author if the manuscript has to be accepted, not be accepted or has to be accepted after major/minor modifications. If a revised manuscript is not received within three months, and is received later, it will be treated as a new manuscript. Manuscripts not accepted for publication shall not be returned. Correction of proofs Proofs of the articles will be sent to the corresponding author for checking its correctness and completeness. No major or any editorial change in the text or data or in the authors shall be allowed at this stage. The authors are responsible for the final accuracy of the proofs of their manuscripts. The corrected proofs must be returned promptly (maximum within five days). The fax or scanned version (as e-mail attachment) of the corrected proofs is also acceptable. A delay in returning corrected proofs will result in their delayed publication or their publication as they stand. The reference section of a manuscript is usually prone to maximum errors, and hence authors are advised to check this section very carefully. Manuscripts and presentation Manuscripts, in English, must be printed (Times New Roman, 12 size; preferably black laser print) on one side of a white, good quality standard size bond paper. All manuscripts must be typed double spaced and the pagination should be done on the top left corner. Only one set of original illustrations and two first-class copies are adequate. The order of presentation should be: title Page with title, authors, affiliation, corresponding author (with full address, telephone and fax number and e-mail ID), Abstract, Introduction, Materials and Methods, Results, Discussion, Acknowledgments and References. Abstract, tables and legends for figures should be typed on separate pages and not in continuation of the main text. At least one inch margin must be available on all four sides of the paper, to provide space for editorial markings/comments. Title: Title of the paper should be short and yet sufficiently descriptive and informative. Authors: The names of authors should include initials and surname and their institutional addresses. Running title: A short running title not exceeding 60 characters including spaces should be provided. Abstract: The abstract should be limited to approximately 250 words and should contain a line of introduction, important results and their significance. In no case it should be just a statement of results only. References are not allowed in the abstract. Keywords: 3-5 keywords arranged in alphabetical order should be provided below the abstract. Introduction: Introduction should provide the essential background, must briefly review the relevant literature, identify the question(s) to be answered and should state the reasons for undertaking the study. Further, to avoid suspense, it should end with a brief statement of results. Material and Methods: The source of the material used should be clearly mentioned. The procedures adopted should be explicitly stated to enable other workers to reproduce the results. New methods may be described in sufficient details, established methods can be just described in brief with appropriate reference(s). Deviations from previously reported methods, if any, should be explained giving the reasons thereof. Results: The results should be as clear as possible and must state only the findings of the study reported. They should be comprehendible to the readers and sequenced in such a way that a logical progression is developed. The data presented in figures and tables should not be repeatedly summarized. Results should not be intermixed with interpretations or conclusions. Discussion: In this section the results should be discussed in a focused way and not just recapitulated. Authors should try to present the principles, generalizations and relationships. They should specifically show how the results and interpretations reported in the manuscript agree or disagree with already published information. The significance and importance of the work must also be discussed sufficiently. Conclusions must be stated clearly and evidence for each conclusion should be summarized. Long, rambling discussion lacking logic and flow should be avoided. Speculations must be avoided to the extent possible. Acknowledgements: Acknowledgements should be brief and made for specific scientific, technological and monetary (grant) assistance. References: List of references should include only published papers and papers in press. Citations of unpublished work, where absolutely necessary, should be incorporated in the text itself (examples: our unpublished observations or Sharma et al. unpublished data or Sharma et al. personal communications). Personal communications must be supported by an authorized letter of permission. References in the text should be cited according to the name and year system (also known as Harvard system) in chronological order, using 'and' for articles with two authors and et al. for those with more than two authors and semi colon between references (Trager and Jensen, 1976; Sharma et al., 1994). References should be listed at the end in alphabetical order and it must be ensured that all references cited in the text are included in the list and vice versa. The following style should be followed: Journal In the list of references at the end of the paper, all the names of authors should be given. However, if there are more than six authors, list first six names followed by et al. Diseases. Houba. V (Ed.), Churchill Living stone, New York. pp 104-129. Conferences Mahajan RC, Malla N, Stella M and Ganguly NK. 1995. Cysticercosis in India: Immunodiagnosis and Treatment. Paper presented at the Twelfth National Congress of Parasitology. Panaji, Goa, India. Reports World Health Organization. 1984. The Leishmaniasis. Technical Report Series No. 701. Geneva, Switzerland. Electronic format papers Ponce LV. 2005. Adoptive transfer of dendritic cells modulates immunogenesis and tolerogenesis in a neonatal model of murine cutaneous leishmaniasis. Kinetoplastid Biology and Disease. 4:2. http://www.kinetoplastids.com/content/4/1/2 . Papers published only with Digital Object Identifier (DOI) numbers Jabakhanji R. Inflammatory and neuropathic pain animals exhibit distinct responses to innocuous and motoric challenges. Molecular Pain. DOI:10.1186/ 1744-8069-2-1. Tables: Tables should be typed separately and numbered consecutively with Roman numerals (I, II, III etc.). Each table should bear a brief title. Column headings should also be brief. Units of measurements should also be abbreviated and placed below the headings. Statistical variations such as SD and SE should be identified. Numbers less than one should have a zero set before the decimal point (e.g. 0.1). Edited books Illustrations: Three sets (one original and two firstclass copies) of illustrations should be submitted, numbered consecutively in Arabic numerals (1, 2, 3 etc.). Line drawings should be made on good quality art paper. Letters, numbers and symbols should be clear in figures and of sufficient size so that when reduced they can be accommodated in a single column (8.5 cm) or double column (17.5 cm) without loss of clarity. Title and explanation of symbols should be typed on separate pages. Data for tables, graphs etc. should be carefully verified. All statistical evaluations, percentage and other calculations should be checked thoroughly before submission of a paper. Once a paper has been accepted for publication, no alterations will be allowed. Schaniz PM and Kagan IG. 1980. Echinococcosis. In: Immunological Investigations of Tropical Parasitic Photographs: Monochrome photographs should be submitted in triplicate (all originals; photocopies are • Trager W and Jensen JB. 1976. Human malaria parasite in continuous culture. Science. 193:673-675. • Cheever AW, Macedonia JG, Mosimann JE and Cheever EA. 1994. Kinetics of egg production and egg excreted by Schistosoma mansoni and S. japonicum in mice infected with a single pair of worms. American Journal of Tropical Medicine and Hygiene. 50:281-285. The name of the journal should either be given in full or abbreviated as per the Index Medicus or Medline. not accepted) and should be un-mounted and numbered consecutively. The title of the paper and author(s), and orientation should be indicated with an arrow should be penciled on the back of each photograph. The photographs should be good quality prints on glossy paper, sharply focused showing good contrast to ensure high quality production. Their size should conform to a single (8.5 cm) or double (17.5 cm) column size. Symbols, arrows and letters should contrast with the background. Colour photographs are also accepted; however, authors will be charged for the same. Legends for illustrations and photographs: Legends for illustrations and captions for photographs should be typed double spaced on a separate page. E n z y m e n o m e n c l a t u re : T h e t r i v i a l n a m e s recommended by the IUPAC-IUB Commissions should be used. At the first citation in the text of the paper, its code number and systematic name should be indicated. Interleukins: Designations recommended by IUIS committee on interleukins have to be used (European Cytokine Network. 1991. vol. 2 (5). pp 309-310). Abbreviations: Only standard abbreviations are to be used. This should conform to the International System of Unit(s), the SI for the Health Professional, WHO, Geneva, 1977. Short communications: These should be prepared in a manner similar to research papers but should not be categorized with the subheadings as in the full paper. However, abstract should be provided. Letters: This is the quickest way of ensuring publications. These should not exceed 500 words. Submission of manuscripts on disk The general style of presentation as detailed above should be followed. Please label your disk with (1) short version of the title of the manuscript, (2) name of the first author, (3) phone and fax number, postal and e-mail address, (4) hard- and software used, including version, (5) and the list of all the file names (eight characters identifying the file before the dot and three characters after the dot for the identification of application). Use only one file for text, references, tables and legends to illustrations. Tables should be placed at the end of text. Use boldface, italics and suband superscript where necessary. Illustrations have to be stored in separate files. Digital figures should be in Word or preferably JPEG files. Preferably, please use MS-Word word-processing package. A compact disk is preferred over a floppy disk. The original and two hard copies with illustrations, containing the exact version on the disk, and the duly complete “Undertaking by Authors” must be submitted along with the disk. JOURNAL OF PARASITIC DISEASES J PD (Official organ of the Indian Society for Parasitology) Undertaking by Authors We, the undersigned, give an undertaking to the following effect with regard to our manuscript titled: _____________________________________________________________________________________________ _____________________________________________________________________________________________ _____________________________________________________________________________________________ submitted for consideration to possible publication in Journal of Parasitic Diseases. 1. Research reported was conducted within the proper animal care and according to the applicable National Laws. 2. The manuscript mentioned above has not been submitted for publication in any form to any other journal. 3. The paper, if published in Journal of Parasitic Diseases, will become the sole copyright of the Indian Society for Parasitology and the article(s) thereof will not be published elsewhere unless written consent is obtained from the Editor-inchief. 4. We also agree to the authorship of the manuscript in the following sequence: 1. 2. 3. 4. 5. 6. 7. 8. 9. 10. Authors' names (in sequence) Signature of authors _________________________________________ _________________________________________ _________________________________________ _________________________________________ _________________________________________ _________________________________________ _________________________________________ _________________________________________ _________________________________________ _________________________________________ _______________________________________ _______________________________________ _______________________________________ _______________________________________ _______________________________________ _______________________________________ _______________________________________ _______________________________________ _______________________________________ _______________________________________ Please note: 1. All authors are required to sign independently in the form and in the sequence given above. A photocopy of this form may also be used. 2. No addition/deletion/or any change in the sequence of the authorship will be permissible at a later stage, without valid reasons. If change is valid , then all authors involved should attest to the change. The decision, however, rests wth the Editor-in-chief. 3. If the authorship is contested at any stage, the manuscript will be either returned or will not be processed for publication. Important In order to facilitate speedy processing of the submitted manuscript, please ensure the following: 1. Manuscripts are prepared as per the format of JPD. 2. Three copies of the manuscript and a brief covering letter. 3. Running title of not more than 60 characters including spaces. 4. Keywords (3-5 words). 5. "Undertaking by Authors", duly complete and signed. 6. Three sets of photographs (if any) with legends and captions on a separate page. 7. A CD containing exactly the hard copy version. The Indian Society for Parasitology Membership Application Name : ____________________________________________________________________________________ Educational Qualification:______________________________________________________________________ Field of Specialization :________________________________________________________________________ Work Address:______________________________________________________________________________ __________________________________________________________________________________________ Phone.:_________________________Fax:____________________________E-mail:______________________ Mailing Address:_____________________________________________________________________________ __________________________________________________________________________________________ Phone.:_________________________Fax:____________________________E-mail:______________________ Signature of the Applicant:_____________________________________________________________________ Proposed by:______________________________ Seconded by:_____________________________________ (Member, ISP) (Member, ISP) __________________________________________________________________________________________ Membership Fee India : Life Membership : Rs. 2000.00 Other Countries : US$ 400.00 (Demand draft should be drawn in favour of Secretary, The Indian Society for Parasitology, Payable at Lucknow) Membership of the Society includes subscription to Journal of Parasitic Diseases, a biannual publication of the ISP. Members are eligible to attend annual conferences of the ISP and avail discounts in registration fee. Membership is governed by the Constitution and Bye Laws of the Indian Society for Parasitology. ----------------------------------------------------------------------------------------------------------------------------------------(For Office use Only) Life Membership No. :____________________ Receipt No.:___________________ Signature of Secretary Signature of Treasurer ---------------------------------------------------------------------------------------------------------------------------------------Note: Duly completed form along with draft should be mailed to Dr. J. K. Saxena Secretary, ISP, Division of biochemistry, Central Drug Research Institute, Post Box-173, Chattar Manzil Palace, Lucknow-226001, India. Personal cheques/cash are not accepted. There is no annual or ordinary membership. This form may be xeroxed. THE INDIAN SOCIETY FOR PARASITOLOGY Executive Committee President Secretary Professor Veena Tandon Department of Zoology North-Eastern Hill University Shillong Dr. J. K. Saxena Division of Biochemistry Central Drug Research Institute Lucknow Vice-President Joint Secretary Dr. J. Mahanta Regional Medical Research Centre Dibrugarh Dr. Vas Dev Malaria Research Center Sonpur Treasurer Dr. L. M. Tripathi Division of Parasitology Central Drug Research Institute Lucknow Members Dr. Wasim Ahmed, Aligarh Dr. P. Prakash Babu, Hyderabad Dr. S. C. Dutta, Kolkata Dr. S. K. Ghosh, Bangalore Dr. Neena Goyal, Lucknow Prof. Neelima Gupta, Bareilley Dr. B. V. Jadhav, Aurangabad Prof. P. D. Juyal, Ludhiana Dr. A. M. Khan, Dibrugarh Dr. Ashwani Kumar, Goa Prof. G. G. Mani, Visakhapatnam Dr. S. K. Puri, Lucknow Prof. R. Kaleysa Raj, Trivandrum Dr. K. K. Saxena, Bareilley Prof. Prati Pal Singh, S. A. S. Nagar Statements and opinions expressed in Journal of Parasitic Diseases or in the presentations during the regular meeting of the Indian Society for Parasitology are those of the author(s) and do not necessarily reflect the official position of the Society. The Editorial Board, Publisher and the Society disclaim any responsibility for the accuracy of statements made by the contributors. Copyright ©2006 The Indian Society for Parasitology Designed & Printed at :- Azad Offset Printers (P) Ltd. 144-Press Site, I.A.-1, Chandigarh Tel. : 0172-2654411, 2652349, 2651316
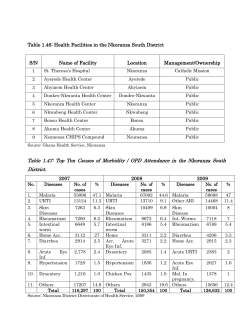









© Copyright 2025